Effekterne af en digitaliseret barndom på hjernens kognitive og følelsesmæssige modning
Illustration: Keren Grafen.
I sin artikel forklarer Dr. Keren Grafen, at mens den menneskelige hjerne indeholder næsten alle nerveceller ved fødslen, sker dens funktionelle modning overvejende postnatalt.
Afgørende processer som synaptogenese, myelinisering og dannelsen af stabile neurale netværk fortsætter ind i den unge voksenalder.
Særligt betydningsfulde er de såkaldte kritiske udviklingsfaser, perioder med øget neuronal plasticitet, hvor miljømæssige påvirkninger i høj grad former hjernens struktur.
I disse faser besluttes det, hvilke synaptiske forbindelser der stabiliseres, og hvilke der igen nedbrydes (synaptisk “eliminering”).
Bevægelse, sanseoplevelser, sprog og social interaktion struktureres. Digitale medier kan ikke erstatte disse oplevelser, da de overvejende er todimensionelle, stillesiddende og socialt reducerede.
Artiklen her var bagrundsmaterialet for den forelæsning, som Dr. Keren Grafen holdt på den årlige konference om miljømedicin i Berlin, 7.-8. november 2025. Artiklen er oversat fra den engelske udgave. Understregninger er tilføjet og enkelte noter er ligeledes tilføjet.
Et kort resume
Dr. Grafen illustrerede i foredraget og her i artiklen, hvordan de digitale medier skader hjerneudviklingen, noget der også er baseret på hendes egen forskning. Et centralt punkt er sensorisk overbelastning. Hurtige billedsekvenser, algoritmisk styrede nyhedsstimuli og permanent tilgængelighed fører til kronisk sensorisk overstimulering og endda til afhængighed. Hippocampus, som er en central struktur for læringsprocesser, hukommelsesdannelse og følelsesmæssig evaluering, er særligt påvirket.
Under en konstant strøm af stimuli mister hippocampus i stigende grad sin filterfunktion. Mekanismer for langvarig potentiering, dvs. stabil forstærkning af læringsrelevante synapser, blokeres. Ny information absorberes stadig, men integreres og lagres mindre effektivt på lang sigt. Forældre oplever det ofte som overfladisk læring, hurtig distraktion eller vanskeligheder med at huske, hvad de har lært på lang sigt.
Motion som en neurobiologisk nødvendighed
Dr. Grafen understreger de motoriske færdigheders centrale rolle i hjerneudviklingen. En stor del af det neuronale kredsløb er direkte eller indirekte forbundet med bevægelse, især via lillehjernen. Bevægelse fremmer ikke kun motoriske færdigheder, men understøtter også kognitive processer, følelsesregulering og indlæringsevne. Overvejende stillesiddende, skærmbaseret fritid reducerer disse vigtige udviklingsimpulser og hæmmer modningen af neurale netværk.
Resumeet er lånt fra en artikel i diagnose:funk om Keren Grafens og Peter Hensingers foredrag.
Europäischen Gesellschaft für klinische Umweltmedizin (EGKU) 23. årlige konference om miljømedicin, Berlin, 7.-8. november 2025
Effekter af en digitaliseret barndom på hjernens kognitive og følelsesmæssige modning
af Keren Grafen
Brain rot/Hjerneforrådnelse. Et udtryk, der lyder provokerende – og alligevel beskriver det med en skræmmende præcision, hvad jeg ser hver dag i min praksis: Et forfald af neurale strukturer udløst af overdreven, algoritmisk styrede digitale stimuli. Ikke underligt, at Oxford Languages valgte netop dette ord som årets ord i 2024.1 Udtrykket refererer til den kognitive og følelsesmæssige svækkelse forårsaget af overdreven skærmbrug, især hos børn.
For første gang i historien vokser børn op permanent og intensivt udsat for digitale stimuli. Smartphones, tablets, sociale netværk – de er ikke længere bare værktøjer. De er digitale livsrum, hvor barndommen finder sted i dag.
Og det spørgsmål, vi alle skal stille os selv, er ikke kun et medicinsk eller uddannelsesmæssigt et, men et socialt et: Hvad sker der med en hjerne, der modnes gennem todimensionelle, algoritmisk styrede stimuli i stedet for gennem direkte oplevelser, berøring, bevægelse og social interaktion? Når likes, hurtige billedsekvenser og swiping dominerer opfattelse, tænkning og følelse, er det ikke kun hjernens arkitektur, der ændrer sig, men også grundlaget for, hvad vi forstår som vores “selv”.
De seneste data fra Common Sense Media-studiet fra 2025 2 giver imponerende tal om den eksplosive stigning i mediebrug blandt småbørn og børn. Børn helt ned til to og fire år bruger allerede over to timer om dagen med skærmmedier. Spilletiden er steget med 65 procent i de sidste fire år mellem 2020 og 2024, og næsten hvert tredje barn bruger AI-baserede applikationer til læring. I en alder af to år har fire ud af ti børn allerede deres egen tablet, og i en alder af fire år er det mere end halvdelen. I en alder af otte år har næsten hvert fjerde barn deres egen smartphone. Korte videoplatforme, oprindeligt designet til ældre børn og teenagere, bliver nu i stigende grad den primære kilde til videoindhold for små børn. Størstedelen af forældrene udtrykker stor bekymring over skærmtiden. Daglig læsning er faldende blandt børn i alderen fem til otte år. Det er også bekymrende, at 20 % af børnene op til otte år allerede bruger digitale enheder til at falde til ro, under måltider eller til at falde i søvn med.
Denne artikel har som formål at give indsigt i vores neurobiologiske forskningsresultater og forklare, hvordan digitale miljøer påvirker hjernens kognitive og følelsesmæssige modning.
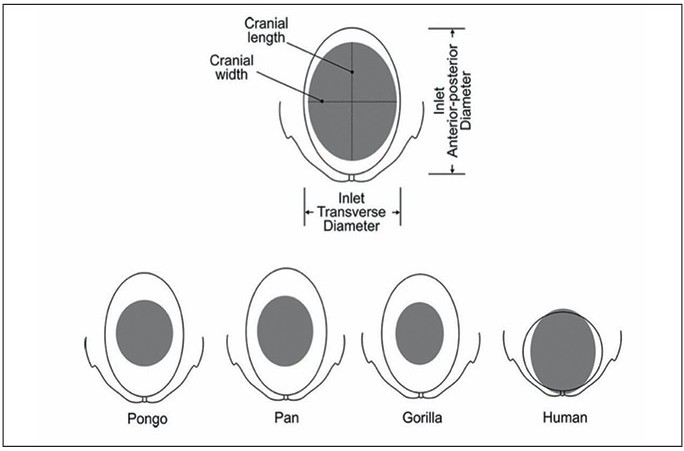
1. Obstetrisk * dilemma: Evolutionært kompromis mellem bækkenstørrelse og hjerneudvikling
Menneskets evolution er karakteriseret ved et bemærkelsesværdigt kompromis mellem bevægelse og hjerneudvikling. Med udviklingen af oprejst gang tilpassede de anatomiske strukturer sig, især dem i kvindens bækken. Et bækken, der var for bredt, ville have kompromitteret stabiliteten ved oprejst gang, mens et, der var for smalt, ville have gjort det vanskeligt at føde børn med relativt store hoveder. Samtidig fortsatte hjernevolumen med at vokse for at muliggøre komplekse kognitive evner såsom tænkning og sprog. 3, 4, 5 Denne konflikt mellem bækkenstørrelse og hovedvolumen førte til det, der er kendt som det obstetriske dilemma. 6 Den evolutionære løsning på dilemmaet var, at menneskebørn blev født, før deres hjerner var fuldt udviklede. Som et resultat heraf er nyfødte relativt umodne sammenlignet med andre primater: Selvom de har næsten alle deres neuroner, sker dannelsen af de funktionelle neurale netværk overvejende efter fødslen. 7
* Red.: “Obstetrik er den kirurgiske specialitet som omhandler behandling af mødre og fostre under graviditet og fødsel..[og] omfatter også sygdomme i barselperioden [..].” (Wikipedia)
2. Fra oprejst gang til finmotorik: 80% af neuronerne til bevægelse
Væksten i hjernestørrelse førte til adskillige tilpasninger, der var tæt forbundet med udviklingen af motoriske færdigheder. Kvantitative analyser viser, at mennesker har omkring 86 milliarder neuroner. 38 Det er slående, at omkring 80% af neuronerne er ansvarlige for motoriske færdigheder og er hovedsageligt koncentreret i lillehjernen. Det høje antal motoriske neuroner understreger den centrale rolle, som bevægelse og sensorimotorisk integration spiller for den menneskelige hjerne. Lillehjernen er primært ansvarlig for koordinering af bevægelse, finmotorik og balance, men samtidig viser dens omfattende forbindelser til andre områder af hjernen, at fysisk aktivitet er afgørende for udviklingen og funktionen af kognitive netværk.
Børn, der sidder foran digitale enheder i lange perioder og har ringe motorisk aktivitet, bruger ikke denne overvejende andel af motoriske neuroner. Som følge heraf går meget af deres neurale potentiale til spilde, hvilket ikke kun begrænser deres motoriske færdigheder, men kan også have negative effekter på eksekutive funktioner *, indlæringsevne og følelsesmæssig regulering.
* “En betegnelse inden for psykologi og psykiatri for en persons evne til problemløsning, planlægning, gennemførelse af opgaver og regulering af adfærd”
Fysisk træning er derfor afgørende for at fremme neural plasticitet, styrke de kognitive færdigheder og sikre udviklingen af en funktionel, adaptiv hjerne.
3. Ungdomsårene som en kritisk fase af kognitiv modning
Hos mennesker er hjernens udvikling ikke fuldført ved fødslen, men fortsætter ind i voksenalderen, cirka indtil 18-20-årsalderen. Det sker meget langsomt (ekstensiv neuromodning), hvor modningsprocesser finder sted langs forskellige gradienter: fra bagfra til forfra (kaudal → rostral), fra central til perifer (kerneområder før kortikale områder) og fra medial til lateral (mediale strukturer før laterale associationsbarker), og på en måde så hver fase fungerer som fundament for den næste. 8, 9
Miljøet og hjernemodningen interagerer gennem hele levetiden. Denne interaktion er særligt intens i de såkaldte kritiske faser. Kritiske faser er udviklingsvinduer, hvor nervesystemet udviser særlig høj plasticitet, og hvor de miljømæssige påvirkninger har en strukturerende effekt. 11-14 I disse perioder reorganiseres og stabiliseres neurale kredsløb funktionelt gennem oplevelsesafhængig forfining, forudsat at de passende sensoriske eller kognitive stimuli er til stede. 10,11 Hvis en kritisk fase afbrydes, eller den nødvendige oplevelse ikke gives, kan det forårsage langvarige og til tider uoprettelige forringelser i udviklingen. 8
Sådanne kritiske perioder omfatter f.eks. tidlig sensorimotorisk modning, hvor grundlæggende motoriske og perceptuelle funktioner udvikles; fasen med motorisk læring, som tjener til at udvikle grov- og finmotorik; og fasen med sprog- og lateraliserings udvikling, som er vigtig for læsning, skrivning og komplekse kognitive evner; modningen af frontallappen (PFC, præfrontal cortex), som muliggør arbejdshukommelse, social adfærd, forventning og begreber om rum og tid; samt den livslange udvikling af det limbiske system, som fungerer som en selektor for nyheder, opmærksomhed, følelser og motivation.
Ifølge Larsen og Luna 15 kan ungdomsårene som helhed (fra fødsel til voksenalder) endda forstås som en storstilet kritisk fase for udviklingen af højere kognitive funktioner.
I disse faser har miljøet en særlig stærk effekt ved at stabilisere de hyppigt anvendte synaptiske forbindelser og eliminere de sjældent anvendte (synaptisk eliminering), hvilket fører til dannelsen af effektive og specialiserede neurale netværk. 11, 13 Ved afslutningen af den kritiske fase falder plasticiteten, hvilket får det indhold, der skal læres, til at blive strukturelt og funktionelt “vævet” ind i hjernen, hvilket skaber stabilitet, men samtidig reducerer fleksibiliteten. 14
Det er værd at bemærke, at kritiske perioder er tidsvinduer, hvor hjernen er særligt formbar, men også særligt følsom. Det skyldes, at i denne periode stabiliseres de neurale forbindelser, som senere danner grundlag for komplekse funktioner. Tidlige, reelle oplevelser – bevægelse, orientering, fysisk kontakt, forskellige sanseindtryk og sociale interaktioner – er afgørende for, at disse netværk kan udvikle sig optimalt. Digitale medier kan kun erstatte denne mangfoldighed i begrænset omfang. Hvis vigtige oplevelser mangler eller formidles på en ensidig måde, kan det permanent forringe udviklingen af centrale hjernefunktioner. Kritiske perioder viser således, hvor fundamental kvaliteten af miljøkontakter er for hjernens udvikling, og hvorfor et aktivt engagement i den virkelige verden fortsat er uerstatteligt.
4. Livslang kritisk fase i hippocampus
Mens den kritiske fase for modningen af mange områder af hjernen kun varer frem til voksenalderen, forbliver den livslang i hippocampus, da der her kontinuerligt dannes nye nerveceller fra et embryonalt kimlag.
Hippocampus er en central struktur i den mediale (dvs. den indre del) af temporallappen, der spiller en nøglerolle i bearbejdningen af hukommelses indhold, rumlig orientering og følelsesmæssig information. Anatomisk set tilhører den den evolutionært ældre archicortex og danner en central komponent i det limbiske system. Dens navn skyldes dens karakteristiske form, som ligner en søhest. I den menneskelige hjerne er den aflang og let buet, med en forreste del pegende fremad og nedad og en bageste del pegende bagud og opad.
Langs denne akse opfylder hippocampus to centrale funktioner, 15 mellem hvilke der er flydende overgange:
- Den anteriore hippocampus er stærkere forbundet med følelsesmæssige processer, herunder social adfærd og angstregulering.
- Den posteriore hippocampus er overvejende forbundet med kognitive opgaver, især rumlig navigation, figurhukommelse (hukommelse af visuel eller billedlig information og mønstre) og deklarativ hukommelse (bevidst genfindbar viden om fakta og begivenheder).
Samtidig skelner hippocampus mellem velkendt og ny information samt fungerer som en selektor for om noget er nyt. Dens evne til at genkende, udvælge, filtrere og integrere relevante signaler i hukommelsesprocesser er baseret på den dynamiske interaktion mellem læringsprocesser koblet med opmærksomhed og motivation. På Institut for Neuroanatomi under professor Dr. Teuchert-Noodt udviklede vi vores model for hippocampus plasticitet og læringsintegration, som forklarer, hvordan disse mekanismer fungerer.
5. En model for de centrale læringsmekanismer i hippocampus
Lad os dykke ned i læringens kerne. Vores model kan opdeles i tre centrale mekanismer, der tilsammen muliggør udvælgelse og integration af nyt læringsindhold.
1. Synaptiske og molekylære mekanismer til læringsforstærkning gennem Hebbiske læringssynapser i hippocampus
Sensorisk information fra omgivelserne når primært hippocampus via den mediale og laterale entorhinale cortex (EC): Den mediale EC koder primært for rumlig information, mens den laterale EC transmitterer objektrelevant information. 20, 21 Disse signaler når granulacellerne i gyrus dentatus, en del af hippocampus, hvor de aktiverer glutamaterge synapser i de øverste to tredjedele af cellelaget. 20, 22 Det er netop her, læring finder sted, som beskrevet af Hebbs læringsregel: “Neuroner, der skyder sammen, forbinder sig sammen”. 22, 23
Det sker molekylært via to typer receptorer – AMPA og NMDA. 23 Når den præsynaptiske neuron bliver aktiv, frigiver den glutamat. Det binder sig initialt til AMPA-receptorerne i den postsynaptiske membran, hvilket får natriumioner (Na+) til at strømme ind i cellen og depolarisere membranen. 23 NMDA-receptoren reagerer også på glutamat, men blokeres initialt af en magnesiumion (Mg2+), der fungerer som en “prop” for at forhindre strømmen af ioner. 23, 24
Kun når selve postsynapsen er aktiv og depolariseret, fjernes Mg2+-blokken, og NMDA-receptoren kan åbne sig. Når glutamat derefter binder sig til den åbne NMDA-receptor, strømmer calcium (Ca2+) ind i postsynapsen. Dette calcium er det afgørende signal for synaptisk amplifikation, kendt som langtidspotentiering (LTP). Præsynapsen og postsynapsen aktiveres næsten samtidigt. NMDA-receptoren fungerer som en “OG-switch” – kun når begge er aktive, styrkes forbindelsen. 23, 24
Der findes to former for LTP: Tidlig LTP styrker synapsen inden for få minutter i flere timer ved at inkorporere AMPA-receptorer. Sen LTP virker på det strukturelle niveau, hvilket fører til ny proteinsyntese og synapsedannelse over timer til dage og sikrer varig styrkelse. 23, 24
Dette system er specifikt rettet mod læring: information fra entorhinal cortex, især rumlig information, styrker specifikt synapserne og konsoliderer læringsprocesser. Hvis denne information mangler, sker der ingen styrkelse, og der finder ingen læring sted. Desuden atrofierer (svinder ind) ubrugte forbindelser – i henhold til princippet om “brug det eller miste det”. 20-24
2. Neurogenese: Permanent rastløshed tvinger reorganisering
Mens sensoriske input i det molekylære lag af gyrus dentatus initialt behandles i de øverste to tredjedele, finder en anden synaptisk forbindelse sted i den nederste tredjedel. Umiddelbart under granulacellelaget ligger et livslangt aktivt embryonalt kimlag – den såkaldte subgranulære zone (SGZ). 25, 26 Her produceres kontinuerligt nye nerveceller: cirka 1600 celler pr. mm² hos børn og omkring 700 celler pr. mm² hos voksne.
For at de nydannede granulaceller kan integreres i det eksisterende netværk, kræves der en kontinuerlig strukturel og funktionel reorganisering gennem synaptiske tilpasninger, især i den indre tredjedel af molekylelaget. 25, 26 I vores model forløber denne proces som følger: Under den indre vækst af nye nerveceller sender de unge neuroner neurotrofiske faktorer ud, der stimulerer nabokredsløb til at forbinde sig med dem. Eksisterende synapser løsriver sig fra deres tidligere partnere og forbinder sig med de nye neuroner. 25, 26 Det fører til en tilstand af konstant dynamisk ustabilitet – en slags “funktionel hvileløshed” – der holder netværket fleksibelt og tilpasningsdygtigt. 25, 26
Denne permanente omstrukturering skaber grundlaget for høj tilpasningsevne: Den muliggør hurtig kodning og evaluering af sensorisk information, men bidrager samtidig til forgængeligheden af hukommelsesspor. 27 Information kan effektivt evalueres og kategoriseres på kort sigt, men forsvinder, hvis den ikke overføres til langtidshukommelsesprocesser. Det svarer til korttidshukommelsens funktion. 27
3. Theta-rytme og koblingen af følelser og kognition gennem bevægelse
Et centralt træk ved hippocampussystemet er dets timing via theta-rytmen (4-7 Hz). Denne rytme genereres af det mediale septum og fungerer som en “neuronal metronom”, der præcist styrer hippocampus’ aktivitet på en faseafhængig måde. 15, 28 Theta-timing koordinerer informationsstrømmen mellem den anteriore hippocampus, som primært er ansvarlig for følelsesmæssige og sociale processer – herunder angstregulering, affektiv evaluering og motivationstilstande – og den posteriore hippocampus, som specialiserer sig i kognitive opgaver såsom rumlig navigation, kontekstuel hukommelse og deklarativ hukommelse. 15, 29, 30
I en hviletilstand er theta-rytmen normalt ikke aktiv. Den aktiveres kun under aktiv bevægelse, udforskning, fokuseret opmærksomhed eller REM-søvn. 28,29 I disse faser synkroniserer theta-timing neural aktivitet på en faseafhængig måde, så læringsindhold kan bearbejdes på en tidsmæssigt og rumligt organiseret måde. Det gør det muligt at kode ny information effektivt og dynamisk koble følelser/motivation og kognition. 15, 28, 29
Sammen danner disse tre mekanismer grundlaget for, at hippocampus fungerer som en central grænseflade til læring. Den modtager ikke blot information passivt, men kontrollerer den konstant for nyheder, filtrerer den specifikt og koordinerer den endda på forskellige tidsniveauer.
Det gør den til en vigtig integrations- og selektionsinstans i hjernen, der sikrer, at læringsprocesser kan opretholdes fleksibelt, adaptivt og gennem hele livet. Hvis rige, multisensoriske oplevelser mangler og erstattes af stærkt visuelle digitale stimuli, kan det hindre dannelsen af stabile hukommelsesspor og forstyrre den følelsesmæssige selvregulering. Teuchert-Noodt beskriver passende denne effekt som “funktionel afkobling af det limbiske system”. 31
6. Overstimulering af hippocampus af digitale medier og konstante sensoriske stimuli
Vore dages allestedsnærværende brug af digitale medier fører til kronisk overstimulering og dermed til en “funktionel afkobling af det limbiske* system”.
* Det limbiske system omfatter basale deler af hjernen og påvirker følelser og hukommelse.
Uendelige reels, konstante notifikationer og multisensoriske stimuli skaber en uafbrudt strøm af information. Ikke kun lys, farve og bevægelse på skærme aktiverer hippocampus, men også den hurtige rækkefølge af visuelle, auditive og andre sensoriske stimuli.
Termiske effekter såsom blåt lys og overophedning, men også atermiske effekter – såsom hurtige billedændringer eller algoritmisk styret sensorisk overbelastning – skaber en tilstand af permanent aktivering. Neurale netværk der er under konstant stress, kan ikke finde ro og mister i stigende grad deres evne til at adskille relevant fra irrelevant information. 32, 33
Studier giver alarmerende resultater: Børn og unge, der i vid udstrækning bruger medier, viser ændringer i områder, der er ansvarlige for følelsesregulering, belønningsbehandling og kognitiv kontrol. 32, 34 Dyremodeller viser, at overdreven sensorisk stimulering (ESS) i den tidlige udviklingsfase fremmer hyperaktivitet og opmærksomhedsunderskud og forstyrrer lærings- og hukommelsesprocesser. 33, 36 Selv små børn, der bruger skærme intensivt, viser betydelige underskud i skrivning og sprog. 32, 34
I forbindelse med vores model betyder det, at hvis hippocampus konstant oversvømmes med ufiltreret, intens sensorisk input – uden reelle, tredimensionelle oplevelsesrum, uden koblede motoriske færdigheder, uden pauser og gentagelsesfaser – så bliver de tre centrale mekanismer ubalancerede:
- Hebbiske læringssynapser kan ikke styrkes selektivt
- Livslang neurogenese mister sin strukturelle effekt
- Theta-rytmesynkronisering forstyrres 33, 35, 36
Resultatet: Systemet overbelaster sig selv. Netværksstabiliteten falder, indlærings- og hukommelsesevnen forringes, og evnen til at bearbejde information samt lære svækkes. 32-36
Vi taler ikke om en abstrakt fare her, men snarere om meget konkrete, målbare neurologiske konsekvenser for børn og unge, hvis hjerner stadig er i kritiske udviklingsstadier. Den konstante digitale overbelastning virker som en kontinuerlig ild på hjernens centrale læringssystem – med konsekvenser, som vi allerede kan se i dag, og hvis langsigtede effekter vi kun kan gisne om. 32-36
| Skærmbrug blandt mindre børn (<alder 8 år) stiger hurtigt: * 2 – 4 år: > 2 timer/dag foran billedskærme * Gaming: 65 % øgning 2020 – 2024 * KI-lærings-app’er: Bruges af hvert 3. barn * Har eget udstyr med skærm: – 2 år: 40 % har egen tablet – 4 år: 58 % har egen tablet – 8 år: hver fjerde har egen smartphone * Korte videoer/reels: Meget populær blandt små børn * Forældre: Mange bekymrer sig, men ved ikke hvordan de skal reagere * Læsning: Daglig læsning blandt 5-8-åringer falder * Brug af skærmudstyr for at holde børn rolige: ca. 20 % Kilde: Common Sense Media. (2025, February 26). The 2025 Common Sense Census: Media use by kids zero to eight. https://www.commonsensemedia.org/research/the-2025-common-sense-census-media-use-by-kids-zero-to-eight |
7. Aktivering af belønningssystemet og indflydelsen på den præfrontale cortex
Permanent overstimulering af hippocampus-systemet har en effekt langt ud over hippocampus og påvirker især den mesokortikale dopaminvej*. Denne projektion er central for udviklingen og funktionen af den præfrontale cortex (PFC), den region, der er ansvarlig for arbejdshukommelse, impulskontrol, beslutningsprocesser og langsigtet planlægning. Forskning fra Teuchert Noodt-arbejdsgruppen har vist, at den mesokortikale dopamin-innervation af PFC er særligt følsom over for miljømæssige påvirkninger under den tidlige udvikling. Selv let dopaminerg overstimulering i følsomme faser kan føre til dybtgående strukturelle ændringer. Resultaterne viser, at en enkelt dosis metamfetamin i det tidlige udviklingsvindue kan reducere den efterfølgende dopaminfibertæthed i PFC med mere end halvtreds procent. 25, 26 Dette resultat illustrerer, hvor sårbart systemet er, og hvor følsomt det er over for tidlig stimulering i barndommen – en mekanisme, der også er relevant for moderne digital overstimulering i billedlig forstand.
* Se https://en.wikipedia.org/wiki/Mesocortical_pathway og https://lex.dk/dopamin for en oversigt over de forskellige veje for distribution af dopamin til hjernen
Den mesokortikale dopaminvej stimuleres dog også permanent i voksenalderen. Den resulterende kontinuerlige frigivelse af dopamin fører til kronisk aktivering af præfrontale netværk. I stedet for en fint moduleret, kontekstafhængig dopamineffekt forekommer overstimulering. Samtidig overtager GABAerge interneuroner* i PFC i stigende grad moduleringen af dopaminerge fibre. Denne reorganisering repræsenterer et forsøg fra systemet på at kompensere for kronisk overstimulering, men fører til dysregulering af det præfrontale netværk. Resultatet er en betydelig forringelse af eksekutive funktioner: arbejdshukommelse, beslutningsevne, impulskontrol og målorienteret planlægning mister stabilitet og effektivitet. Hjernen reagerer mere og mere impulsivt, søger nye stimuli hurtigere og mister evnen til at opretholde opmærksomheden over længere perioder.
* “Interneuroner er nerveceller som forbinder to nærliggende nerveceller (neuroner) med hinanden. […] Interneuronene udskiller neurotransmitteren GABA og kan dermed virke hæmmende på tilkoblede projektionsneuroner.”(https://sml.snl.no/internevroner)
Samlet set tyder de neurobiologiske fund på, at konstant digital stimulering er en form for kronisk dopaminerg stress, der har en særlig dybtgående effekt på funktionen af den præfrontale cortex. Da dette område fortsætter med at udvikle sig langt ind i den unge voksenalder, påvirker sådanne forstyrrende faktorer et system, der forbliver formbart og sårbart i årevis. Den permanente aktivering af belønningssystemet destabiliserer således ikke blot hippocampus, men svækker også på lang sigt de præfrontale funktioner, der er uerstattelige for selvkontrol, reflekterende tænkning og bæredygtig læring.
Konklusion – Digital kokain for hjernen
For vores børn er den konstante sensoriske overbelastning fra digitale medier som en neurobiologisk spærreild, en form for “digital kokain”. Hvert klik, hvert nyt billede, hvert like udløser små dopamin-kick – belønningssystemet er konstant i højeste alarmberedskab. Hippocampus arbejder uophørligt, den præfrontale cortex er under konstant stress.
Hjernen lærer kun at reagere på hurtige klik – i stedet for dybt forankret, bæredygtig læring. Koncentration, tålmodighed, langsigtet målforfølgelse – alt dette svækkes. Følelser og motivation bliver i stigende grad afkoblet fra kognition og refleksion.
Hvad der er tilbage, er et system, der konstant kræver nye stimuli, men finder mindre og mindre reel tilfredsstillelse. Digital overstimulering er ingen erstatning for reel oplevelse, berøring eller social interaktion. Den er normalt bevægelsesfri, hvilket gør omkring 80% af hjernen, som er beregnet til motoriske funktioner, praktisk talt ubrugelig. Hjernen bliver således systematisk frarøvet sine naturlige lærings- og handlingsevner. Kort sagt, den konstante overstimulering i den digitale verden sætter vores hjerner i en kunstig tilstand – afhængig af nyheder, men mangler dybde.
Ansvar og mulighed
Vi bærer alle et ansvar for det: som forældre, lærere, forskere, læger og terapeuter. Den særligt lange postnatale hjerneudvikling – på grund af kompromiset mellem bækkenbredde og hjernestørrelse – samt den uforholdsmæssigt høje tæthed af motoriske neuroner i hjernen (80%) nødvendiggør reelle, tredimensionelle oplevelser: Bevægelse, forståelse, følelse og social nærhed. Kun på den måde kan der udvikles stabile kortikale og limbiske netværk, som danner grundlag for kognitive, følelsesmæssige og sociale færdigheder.
Ligesom når vi håndterer alkohol eller kører bil, har vi brug for regler, strukturer og beskyttende foranstaltninger for at forhindre vores børns hjerner i at rådne – for at forhindre hjerneråd i at blive normen.
Det er op til os at undersøge de digitale miljøer kritisk, anerkende algoritmisk kontrol og skabe betingelser, hvor børn kan vokse op og blive robuste, selvbestemmende og empatiske.
Vi står ved en skillevej: Vi kan forme og kontrollere risiciene i den digitale verden, eller vi kan overlade vores børn til en endeløs strøm af stimuli.
Digitale medier kan i betydelig grad forringe børns kognitive og følelsesmæssige udvikling. Vi skal aktivt støtte vores børn, give dem vejledning og navigere dem sikkert gennem den digitalt drevne verden, så deres hjerner kan udvikle sig til deres fulde potentiale.
Der er ingen plan B. Der er kun denne ene barndom. Lad os beskytte den. For det, vi bevarer i dag, bestemmer, hvem vores børn vil være i morgen.
Denne artikel blev præsenteret som et foredrag på den 23. årlige konference om miljømedicin i European Society for Clinical Environmental Medicine (EGKU).

Forfatter:
Dr. rer. Nat. Keren Grafen
Neurobiolog siden 2013 med egen klinik.
Studerede biologi, forskede og opnåede sin doktorgrad ved Bielefeld Universitet, hvor hun var professor i neuroanatomi og professor i kognitiv neurovidenskab.
E-Mail: info@praxis-biomed.de
Litteratur
1. Oxford University Press. Oxford University Press; [(accessed on December 2, 2024)]. Brain Rot Named
Oxford Word of the Year 2024. Available online: https://corp.oup.com/news/brain-rot-named- oxford
word-of-the-year-2024/
2. Common Sense Media (2025). The Common Sense Census: Media Use by Kids Age Zero to Eight 2025 Common Sense Media. Available online: https://www.commonsensemedia.org/sites/default/files/
research/report/2025-common-sense-census- web-2.pdf
3. Rosenberg, K., & Trevathan, W. (2005). Bipedalism and human birth: The obstetrical dilemma revisited.
Evolutionary Anthropology: Issues, News, and Reviews, 14(5), 161–168. https://doi.org/10.1002/
evan.1360040506
4. Washburn, S. L. (1960). Tools and human evolution. Scientific American, 203(3), 63–75.
5. Kuzawa, C. W., Chugani, H. T., Grossman, L. I., Lipovich, L., Muzik, O., Hof, P. R., Wildman, D. E.,
Sherwood, C. C., Leonard, W. R., & Lange, N. (2014). Metabolic costs and evolutionary implications of
human brain development. Proceedings of the National Academy of Sciences of the United States of
America, 111(36), 13010–13015. https://doi.org/10.1073/pnas.1323099111
6. Wittman, A. B., & Wall, L. L. (2007). The evolutionary origins of obstructed labor: Bipedalism, encephal
ization, and the human obstetric dilemma. Obstetrics & Gynecology Survey, 62(11), 739–748. https://
doi.org/10.1097/01.ogx.0000286584.04310.5c
7. Teuchert-Noodt, G. (2016). The development of the child’s brain is subject to the triad of activity,
dynamics, and compensation. Trauma – Journal for Psychotraumatology and its Applications, 14(2).
8 Hensch, T. K. (2004). Critical period regulation. Annual Review of Neuroscience, 27, 549–579. https://
doi.org/10.1146/annurev.neuro.27.070203.144327
9. Knudsen, E. I. (2004). Sensitive periods in the development of the brain and behavior. Cognitive
Neuroscience, 16(8), 1412–1425. https://doi.org/10.1162/0898929042304796
10. Levelt, C. N., & Hübener, M. (2012). Critical-period plasticity in the visual cortex. Annual Review of
Neuroscience, 35, 309–330. https://doi.org/10.1146/annurev-neuro-061010-113813
11. Hooks, B. M., & Chen, C. (2020). Circuitry underlying experience-dependent plasticity in the mouse
visual system. Neuron, 106(1), 21–36. https://doi.org/10.1016/j.neuron.2020.01.031
12. Petanjek, Z., et al. (2011). Extraordinary neoteny of synaptic spines in the human prefrontal cortex.
Proceedings of the National Academy of Sciences, 108(32), 13281–13286.
13. Takesian, A. E., & Hensch, T. K. (2013). Balancing plasticity/stability across brain development. Progress
in Brain Research, 207, 3–34.
14. Larsen, B., & Luna, B. (2018). Adolescence as a neurobiological critical period for the develop
ment of higher-order cognition. Neuroscience & Biobehavioral Reviews, 94, 179–195. https://doi.
org/10.1016/j.neubiorev.2018.09.005
15. Strange, B. A., Witter, M. P., Lein, E. S., & Moser, E. I. (2014). Functional organization of the hippocam
pal longitudinal axis. Nature Reviews Neuroscience, 15(10), 655–669. https://doi.org/10.1038/
nrn3785
16. Strange, B. A., Fletcher, P. C., Henson, R. N., Friston, K. J., & Dolan, R. J. (1999). Segregating the
functions of human hippocampus. Proceedings of the National Academy of Sciences of the USA, 96(7),
4034–4039. https://doi.org/10.1073/pnas.96.7.4034
17. Kaplan, R., Horner, A. J., Bandettini, P. A., Doeller, C. F., & Burgess, N. (2014). Human hippocampal
processing of environmental novelty during spatial navigation. Hippocampus, 24(7), 740–750. https://
doi.org/10.1002/hipo.22264
18. Tacikowski, P., Kalender, G., Ciliberti, D., & Fried, I. (2024). Human hippocampal and entorhinal neurons
encode the temporal structure of experience. Nature, 635(8037), 160–167. https://doi.org/10.1038/
s41586-024-07973-1
19. Gómez-Ocádiz, R., Trippa, M., Zhang, C.-L., Posani, L., Cocco, S., Monasson, R., & Schmidt-Hieber,
C. (2022). A synaptic signal for novelty processing in the hippocampus. Nature Communications, 13(1), https://doi.org/10.1038/s41467-022-31775-6
20. Strauch, C., Böge, J., Shchyglo, O., Dubovyk, V., & Manahan-Vaughan, D. (2025). The suprapyramidal and infrapyramidal blades of the dentate gyrus exhibit different GluN subunit content and dissimilar frequency-dependent synaptic plasticity in vivo. Hippocampus, 35(2), e70002. https://doi.org/10.1002/hipo.70002
21. Wilson, D. I. G., Watanabe, S., Milner, H., & Ainge, J. A. (2013). Lateral entorhinal cortex is necessary for
associative but not nonassociative recognition memory. Hippocampus, 23(12), 1280– 1290. https://
doi.org/10.1002/hipo.22165
22. Lee, I., & Kesner, R. P. (2002). Differential contribution of NMDA receptors in hippocampal subregions
to spatial working memory. Nature Neuroscience, 5(2), 162–168. https://doi.org/10.1038/nn790
23. Mikulasch, F. A., Rudelt, L., & Priesemann, V. (2021). Local dendritic balance enables learning of
efficient representations in networks of spiking neurons. Proceedings of the National Academy of
Sciences of the United States of America, 118(50), e2021925118. https://doi.org/10.1073/
pnas.2021925118
24. Yamaguchi, M., & Mori, K. (2005). Critical period for sensory experience-dependent survival of newly
generated granule cells in the adult mouse olfactory bulb. Proceedings of the National Academy
of Sciences of the United States of America, 102(27), 9697–9702. https://doi.org/10.1073/
pnas.0406082102
25. Dawirs, R., Hildebrandt, K., & Teuchert-Noodt, G. (1998). Adult treatment with haloperidol increases
dentate granule cell proliferation in the gerbil hippocampus. Journal of Neural Transmission, 105(3–4),
317–327. https://doi.org/10.1007/s007020050061
26. Hildebrandt, K., Teuchert-Noodt, G., & Dawirs, R. (1999). A single neonatal dose of methamphetamine
affects granule cell proliferation in the gerbil hippocampus. Journal of Neural Transmission, 106(7–8),
689–701. https://doi.org/10.1007/s007020050061
27. Zhao, C., Teng, E. M., Summers, R. G., Ming, G.-L., & Gage, F. H. (2006). Distinct morphological stages
of dentate granule neuron maturation in the adult mouse hippocampus. The Journal of Neuroscience,
26(1), 3–11. https://doi.org/10.1523/JNEUROSCI.3648-05.2006
28. Buzsáki, G. (2002). Theta oscillations in the hippocampus. Neuron, 33(3), 325–340. https://doi.
org/10.1016/S0896-6273(02)00586-X
29. Colgin, L. L. (2013). Mechanisms and functions of theta rhythms. Annual Review of Neuroscience, 36,
295–312. https://doi.org/10.1146/annurev-neuro-062012-170330
30. Eichenbaum, H. (2017). Memory: Organization and control. Annual Review of Psychology, 68, 19– 45.
https://doi.org/10.1146/annurev-psych-010416-044131
31. Teuchert-Noodt, G. (2018). Risks of neuroplastic adaptation of the perception of space and time
in the context of media effectiveness. In: J. Weinzirl, P.Lutzka, P.Heusser (Eds.), Significance and
endangerment of the senses in the digital age (Witten Colloquium 5), Würzburg 2017. https://www.
erziehungskunst.de/artikel/das-wird-eine-ganze-generation-in-die-steinzeit-zurueckwerfen
32. Twenge, J. M., Joiner, T. E., Rogers, M. L., & Martin, G. N. (2018). Increases in depressive
symptoms, suicide-related outcomes, and suicide rates among U.S. adolescents after 2010 and
links to increased new media screen time. Clinical Psychological Science, 6(1), 3–17. https://doi.
org/10.1177/2167702617723376
33. Hutton, J. S., Dudley, J., Horowitz-Kraus, T., DeWitt, T., & Holland, S. K. (2020). Associations between
screen-based media use and brain white matter integrity in preschool-aged children. JAMA Pediatrics,
174(1), e193869. https://doi.org/10.1001/jamapediatrics.2019.3869
34. Christakis, D. A., Ramirez, J. S. B., Ferguson, S. M., & Ramirez, J.-M. (2018). How early media expo
sure may affect cognitive function: A review of results from observations in humans and experiments
in mice. Proceedings of the National Academy of Sciences, 115(40), 9851–9858. https://doi.
org/10.1073/pnas.171154811
35. Orben, A., & Przybylski, A. (2019). Screens, Teens, and Psychological Well-Being: Evidence
From Three Time-Use-Diary Studies. Psychological Science. 30. 095679761983032.
10.1177/0956797619830329.
36. Volkow, N. D., Wang, G.-J., Fowler, J. S., Tomasi, D., & Telang, F. (2011). Addiction: Beyond dopamine
reward circuitry. Proceedings of the National Academy of Sciences, 108(37), 15037–15042. https://
doi.org/10.1073/pnas.1010654108
37. Owens, J., Adolescent Sleep Working Group, & Committee on Adolescence. (2014). Insufficient sleep in
adolescents: Causes, consequences, and treatment strategies. Pediatrics, 134(3), e921– e932. https://
doi.org/10.1542/peds.2014-1696
38. Prati, J. M., Pontes-Silva, A., & Gianlorenço, A. C. L. (2024). The cerebellum and its connections to
other brain structures involved in motor and non-motor functions: A comprehensive review. Behavioural
Brain Research, 465,114933. https://doi.org/10.1016/j.bbr.2024.114933