Review og evidenskort over radiofrekvent felteksponering og DNA skader
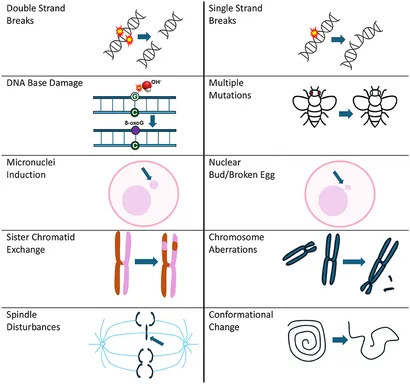
Illustration: Figur 1 fra forskningsartiklen. Typer af DNA-skader rapporteret på grund af RF-eksponering.
På baggrund af evidensbaseret forskning om moderne trådløse teknologier opfordrer forskerverdenen myndigheder og regeringer til at opdatere sikkerhedsforanstaltningerne.
Presset fra forskerne vil kun vokse efterhånden som risiciene i den virkelige verden bliver tydeligere og tydeligere.
En artikel om trådløs radiofrekvent stråling og beskadigelse af DNA, er nylig blevet offentliggjort af forskerne ved Oceania Radiofrequency Scientific Advisory Association (ORSAA).
“…. mellemlang til langvarig RF-EMF-eksponering, især ved lave intensiteter, kan forårsage genetisk skade gennem ikke-termiske mekanismer såsom øget produktion af frie radikaler og oxidativ stress.
Genetisk skade kan have vidtrækkende, langsigtede og potentielt irreversible konsekvenser for individuelle organismer og bredere økologisk og planetarisk sundhed.”
Weller et al. (2025) konkluderer:
“Den samlede evidens viser, at RF-EMF-eksponeringer kan være genotoksiske og kan udgøre en kræftrisiko. Eksponeringens varighed og signaler fra den virkelige verden er de vigtigste faktorer, der påvirker genotoksiciteten… Disse resultater understøtter vedtagelsen af forsigtighedsforanstaltninger sammen med eksisterende termisk-baserede retningslinjer for eksponering.”
Forskningsartiklen Steven Weller et al.: “A scoping review and evidence map of radiofrequency field exposure and genotoxicity: assessing in vivo, in vitro, and epidemiological data.” blev offentliggjort den 30. juli 2025 på Frontiers in Public Health.
Efter en korte introduktion får du hele artiklen oversat til dansk. Den er grundig, men skrevet på forståeligt sprog. Understregninger er tilføjet.
Det nye omfattende review af Weller et al. (2025) vender den tidligere sikkerhedslogik for officiel vurdering af mobiltelefoni på hovedet. Selvom de internationale ICNIRP-grænsevædier næsten udelukkende er baseret på beskyttelse mod vævsopvarmning, viser reviewet af Weller et al. (2025), at stråling fra mobiltelefoner kan have langt større effekt: Over 500 studier fandt overvejende betydelig DNA-skader – ofte endda under de nuværende grænseværdier. Især mobiltelefonsignaler fra den virkelige verden og længere eksponeringstider fører til udtalte effekter. Studiet fokuserer på oxidativ stress som en nøglemekanisme, der gør reproduktions- og hjerneceller særligt sårbare.
De vigtigste resultater af Weller et al. studiet (2025):
- >500 studier evalueret
- 59% viser DNA-skader fra mobiltelefonstråling
- 86% af studierne om basalskade finder effekter
- >50% af effekterne forekommer under ICNIRP-grænseværdierne
- Reelle mobiltelefonsignaler er stærkere end signalgeneratorer
- U-formet dosis-responskurve: meget korte og meget lange eksponeringer er særligt effektive
- Særligt følsomme: sædceller, æggestokkeceller, hjerneceller, insekter, planter, fugle
- Hovedmekanismer: oxidativ stress (83%), forstyrrelse af spindelapparatet (proteinstruktur)
- Brancherelaterede studier: >80% ingen effekter
- Uafhængige studier: ~74% med effekter
- Anbefaling: Revider grænsevædierne, forsigtighedsprincip, uafhængig forskning
Fra diagnose:funk.
Forskningsartiklen
Et undersøgende review og evidenskort over radiofrekvent felteksponering og genotoksicitet: vurdering af in vivo, in vitro og epidemiologiske data
Weller SG, McCredden JE, Leach V, Chu C, Lam AK-Y (2025) A scoping review and evidence map of radiofrequency field exposure and genotoxicity: assessing in vivo, in vitro, and epidemiological data. Front. Public Health 13:1613353. doi: 10.3389/fpubh.2025.1613353
Open Access: https://www.frontiersin.org/journals/public-health/articles/10.3389/fpubh.2025.1613353
Abstrakt
Baggrund
Studier der undersøger genotoksiske effekter af eksponering for radiofrekvente elektromagnetiske felter (RF-EMF) (3 kHz-300 GHz), har brugt en lang række parametre, og resultaterne har været inkonsistente. En systematisk kortlægning af eksisterende forskning er nødvendig for at identificere nye mønstre og for at informere fremtidig forskning og politik.
Metoder
Evidenskortlægning blev udført ved hjælp af vejledning fra Preferred Reporting Items for Systematic reviews and Meta-Analyses for Scoping Reviews (PRISMA-ScR). En omfattende søgestrategi blev anvendt på tværs af flere forskningsdatabaser med specifikke inklusions- og eksklusionskriterier inden for hvert vidensdomæne. Kvantitativ aggregering ved hjælp af tabeller, grafer og varmekort blev brugt til at syntetisere data i henhold til studietype, organismetype, eksponeringsniveau og varighed, biologiske markører (genotoksicitet, cellulær stress, apoptose), RF-EMF-signalkarakteristika samt finansieringskilde for yderligere at kontekstualisere evidenslandskabet. Kvalitetskriterier blev anvendt som en del af en fokuseret analyse for at undersøge potentielle bias og deres effekter på resultater.
Resultater
Over 500 relevante studier blev identificeret, kategoriseret som in vitro (53%), in vivo (37%) og epidemiologiske (10%) og grupperet efter type DNA-skade, organisme, intensitet, varighed, signalkarakteristika, biologiske markører og finansieringskilde. In vitro-studier viste overvejende proportionalt færre signifikante effekter, mens in vivo og epidemiologiske studier viste flere. Studier af DNA-baseskader viste den højeste andel af effekter, ligesom studier, der brugte GSM-taletilstand, pulserede signaler og enheder i den virkelige verden. Der blev identificeret en kompleks sammenhæng mellem eksponeringsintensitet og varighed, hvor varighed fremkom som en kritisk faktor for resultaterne. En kompleks U-formet dosis-respons-sammenhæng var tydelig, hvilket tyder på adaptive cellulære reaktioner med øget produktion af frie radikaler som en plausibel mekanisme. Studier af højere kvalitet viste færre signifikante effekter; finansieringskilden havde dog en stærkere indflydelse på resultaterne end studies kvalitet. Over halvdelen (58%) af de studier, der observerede DNA-skader, anvendte eksponeringer under grænseværdierne fra International Commission of Non-Ionizing Radiation Protection (ICNIRP).
Konklusion
Den samlede evidens viser, at RF-EMF-eksponeringer kan være genotoksiske og kan udgøre en kræftrisiko. Eksponeringens varighed og signaler fra den virkelige verden er de vigtigste faktorer, der påvirker genotoksiciteten, hvilket berettiger til yderligere fokuseret forskning. For at imødegå potentielle genotoksiske risici understøtter disse resultater vedtagelsen af forsigtighedsforanstaltninger sammen med eksisterende termisk-baserede retningslinjer for eksponering.
Introduktion
Baggrund og begrundelse
Verden står over for en kræftpandemi med eksponentiel vækst i mange kræftformer (International Agency for Research on Cancer (IARC) World Cancer Reports) (1–4). Selv om befolkningens aldring bidrager til denne tendens (5), kan den ikke fuldt ud forklare den observerede stigning i visse kræftformer, hvilket tyder på, at miljø- og/eller livsstilsfaktorer spiller en rolle. En potentiel faktor, der skal tages i betragtning, er de stigende globale baggrundsniveauer af menneskeskabte radiofrekvente elektromagnetiske felter (RF-EMF) (6), som falder sammen med en generel stigning i kræftforekomsten i løbet af de sidste årtier. Selvom denne tidsmæssige overlapning ikke indebærer årsagssammenhæng, fremhæver den behovet for omhyggelig undersøgelse af den potentielle rolle af RF-EMF-eksponering blandt de mange miljø- og livsstilsfaktorer, der er forbundet med kræft.
IARC’s nuværende perspektiv er, at RF-EMF-eksponeringer, herunder eksponering for mobiltelefoner, er et muligt kræftfremkaldende stof i gruppe 2B (IARC, maj 2011) (7, 8). Denne klassificering var baseret på den tilgængelige evidens på det tidspunkt, som omfattede både epidemiologiske og dyreeksperimentelle studier. Selv om evidensen blev anset for troværdig, kunne partiskhed og forvirring ikke helt udelukkes (8). En begrænset forståelse af de underliggende mekanismer, der af nogle betragtes som svage (9) forhindrede også en højere klassificering.
Genetisk ændring er en veletableret udløser for kræftudvikling (10). Genotoksicitet er et fysisk eller kemisk agens’ evne til at fremkalde genetisk skade, hvilket kan resultere i genetiske mutationer (11) og udgør en kritisk vej til kræft. Derfor, hvis RF-EMF-eksponering er forbundet med genotoksicitet, vil dette give stærk evidens for en plausibel mekanisme, der beskriver, hvordan RF-EMF kan initiere carcinogenese hos mennesker og potentielt i alle levende organismer. Med den anbefalede prioritering af en IARC (12) vurdering af det kræftfremkaldende potentiale ved eksponering for radiofrekvens, er en upartisk syntese af dokumentationen for RF-induceret genotoksicitet en afgørende ressource, der er nødvendig for en sådan undersøgelse.
Dette review har til formål at afgøre, om RF-EMF-eksponering kan skade DNA og derved potentielt bidrage til den stigende globale forekomst af kræft.
Radiofrekvente eksponeringer
Radiofrekvente elektromagnetiske felter defineres som ikke-ioniserende elektromagnetiske frekvenser i området 3 kilohertz (kHz) til 300 gigahertz (GHz) (13), som befinder sig mellem ekstremt lavfrekvente felter (ELF), dvs. elektriske effektfrekvenser, og infrarødt lys i det elektromagnetiske spektrum. RF-EMF produceres både naturligt (som kosmisk baggrundsstråling, lyn og anden atmosfærisk aktivitet) og fra en lang række menneskeskabte kilder, herunder radar, radio/tv-udsendelsesantenne, satellitkommunikation, mobiltelefonbasestationer, smart meters, smartphones, Bluetooth-enheder, spillekonsoller, babyalarmer, computere, Wi-Fi-routere, mikrobølgeovne, radiofrekvente implanterede enheder, diatermimaskiner og trådløse strømtransmissionsenheder.
Den primære kilde til naturlig, isotrop mikrobølge baggrundsstråling på Jorden er den kosmiske mikrobølgebaggrund (14). Denne stråling falder til jorden på en ikke-polariseret og kontinuerlig måde ved meget lave intensitetsniveauer så lave som 10-18 W/m2 (6) for frekvenser, der anvendes i mobiltelefonens RF-EMF-spektrum. I modsætning hertil er menneskeskabt RF-EMF-eksponering typisk polariseret og pulseret med intensiteter på op til 10 W/m2 for visse frekvenser, der bruges i trådløs kommunikation, som tilladt af Den Internationale Kommission for Beskyttelse mod Ikke-Ioniserende Stråling (ICNIRP) (15) for eksponeringer i offentligheden. Disse vigtige egenskaber adskiller menneskeskabte og naturlige strålingskilder. Biologisk liv har aldrig før oplevet denne type kunstig stråling, som kan have unikke biologiske effekter (16). Radiofrekvente bølger kan trænge ind i menneskelig hud, afhængigt af bølgefrekvens og vævsegenskaber (17). Lavere frekvente RF-bølger trænger dybere ind og når indre organer, mens højere frekvenser lettere absorberes af hudlagene eller reflekteres (18). Trådløse kommunikationsteknologier, såsom mobiltelefoner og Wi-Fi-routere, er afhængige af RF-frekvenser, der kan passere gennem barrierer som vægge, vinduer og tage for at opretholde signaldækningen.
DNA-skader, hvordan det måles
DNA-skader omfatter forskellige former (se Figur 1), herunder enkeltstrengede brud (SSB’er), dobbeltstrengsbrud (DSB’er), kromosomafvigelser, søsterkromatidudvekslinger og tilstedeværelsen af mikrokerner (11), som alle kan påvirke cellulær integritet betydeligt.
Supplerende tabel 1.
Cirka 10.000 DNA-modifikationer sker hver time pr. celle (19), og af disse er SSB’er de hyppigste typer DNA-læsioner (75 %) (19). SSB’er kan opstå ved eksponering for frie radikaler såsom reaktive oxygen- og nitrogenarter (ROS/RNS) (20). SSB’er kan også forekomme som mellemprodukter af DNA-reparationsprocessen eller som et resultat af abortive cellulære enzymaktiviteter (20). Hvis de ikke repareres hurtigt og korrekt, kan SSB’er forstyrre kritiske processer såsom DNA-replikation og transkription, hvilket i sidste ende kompromitterer genomets stabilitet (21). DSB’er, selvom de er sjældnere, er særligt alvorlige, fordi de involverer brud i begge DNA-strenge. DSB’er udgør en betydelig udfordring for reparationsmekanismer og øger risikoen for kromosomafvigelser og mikrokerner, der fungerer som biomarkører for genomustabilitet (22). Akkumulering af ureparerede DNA-skader, herunder SSB’er, DSB’er og tilhørende abnormiteter, har været stærkt impliceret i udviklingen af kræft (11), aldringsrelaterede lidelser (23) og neurodegenerative sygdomme (24).
Hver type DNA-skade har specifikke analysemetoder (f.eks. kometanalyse) til at opdage og evaluere skaden. De typer af DNA-skader og deres tilknyttede analyser, der undersøges som en del af evidenskortet, er beskrevet i Supplerende tabel 1.1
DNA-skader målt via et assay repræsenterer nettoresultatet af tre faktorer: (i) den skade, der induceres ved eksponering for et eksternt stof, der testes, (ii) baseline-niveauet af endogen DNA-skade, der opstår naturligt under normale fysiologiske forhold, og (iii) omfanget af skader, der repareres af de cellulære DNA-reparationsmekanismer.
Den nuværende viden og usikkerheder
Det er veletableret, at de biologiske og sundhedsmæssige effekter af mikrobølger afhænger af forskellige biologiske og fysiske parametre, der adskiller sig på tværs af studier, hvilket fører til variationer i observerede effektresultater (25, 26).
Hundredvis af eksperimentelle studier af varierende kvalitet er blevet udført over mange årtier, der undersøger, om RF-EMF-eksponering kan skade DNA eller resultere i genetiske mutationer. Der er også udført flere reviews, hver med en mindre undergruppe af studier, med blandede resultater. Den balance mellem evidens, der er fastlagt af disse tidligere reviews, spænder fra betydelig evidens for genetisk skade eller interferens, Lai (27), til en smule favoriserende DNA-skade, Ruediger (28), til påstande om, at de fleste studier ikke viser nogen signifikante effekter, Vijayalaxmi og Prihoda (29, 30) og for nylig Romeo et al. (31).
Årsagerne til de store uoverensstemmelser i tidligere reviews omfatter:
1. Omfanget af revisionen, f.eks. kun at fokusere på in vitro-studier eller at dække et mere omfattende sæt in vivo-studier og epidemiologiske studier
2. Den endelige pulje af studier, der udvælges til review (med henblik på udvælgelsesbias)
3. Kvalitetskriterier, der anvendes til at udelukke eller nedklassificere relevante dokumenter.
4. Gennemgå forfatter(e) personlige fordomme eller tilhørsforhold.
I betragtning af den nuværende ufuldstændighed af tilgængelige systematiske oversigter, uoverensstemmelser i rapporteringen og potentielle skævheder er en evidenskortlægning, der undersøger de potentielle genotoksiske effekter af RF-EMF, både rettidig og afgørende.
Formål, anvendelsesområde og mål
Evidenskortet, der præsenteres her, har til formål at foretage en omfattende vurdering af den tilgængelige forskning, der undersøger potentielle genotoksiske effekter forbundet med radiofrekvente (RF) elektromagnetiske felter (3 kHz til 300 GHz) eksponering. Det ultimative mål er at afklare, om RF-eksponering har en plausibel rolle i at skade DNA, med efterfølgende konsekvenser for biologisk sundhed og induktion af kræft.
Omfanget af dette studie omfatter alle større former for DNA-skader såvel som potentielle mekanistiske veje. Studiet undersøger eksperimentel og observationsforskning, herunder in vitro, in vivo og epidemiologiske studier. Den undersøger alle relevante tidligere studier, herunder dem, der er offentliggjort frem til maj 2023. De primære mål er at katalogisere og syntetisere denne evidens, hvilket gør det muligt at identificere potentielle biologiske mekanismer, mønstre, huller og metodologiske begrænsninger på området. Dette studie har til formål at:
• Forstå mangfoldigheden af evidens: At giv et struktureret overblik over de forskellige former for DNA-skader, der er undersøgt i forbindelse med forskellige egenskaber ved RF-eksponering;
• Opdag potentielle biologiske mekanismer: Undersøg og kategoriser de vigtigste biologiske veje og processer, hvorigennem RF-eksponering kan inducere genetisk skade, herunder direkte og indirekte effekter;
• Fremhæve kvalitetsproblemer og skævheder: Identificere styrker og begrænsninger ved eksisterende forskning, herunder metodologisk robusthed, potentielle interessekonflikter og finansieringskilder;
• Bygge bro over datahuller for politik og forskning: Tilbyde indsigt i områder, der kræver yderligere udforskning for at informere fremtidige eksperimentelle design og retningslinjer for folkesundhed;
• Sætte scenen for kvalitetsfokuseret syntese: Lægge grunden til en fremtidig systematisk review og narrativ analyse af studier af høj kvalitet;
• Undersøge eksponeringsniveauer i studier under ICNIRP’s erhvervsmæssige grænse for lokaliserede eksponeringer: Bestem, om de nuværende internationale sikkerhedsretningslinjer er effektive til at beskytte alle levende enheder mod genetiske skader forbundet med RF-eksponeringer.
Primære spørgsmålspunkter
Det primære spørgsmål for dette afgrænsningsreview og evidenskort er: Hvilken evidens findes der for det genotoksiske potentiale af menneskeskabte RF-EMF-eksponeringer? En systematisk kortlægning og syntese af følgende faktorer vil hjælpe med at løse dette spørgsmål:
1. forskningspublikationers placering og historie
2. organismer og celletyper til vurdering af differentiel følsomhed over for radiofrekvenseksponering
3. balance mellem evidens på tværs af in vitro-, in vivo– og epidemiologiske studier for forskellige typer DNA-skader
4. eksponeringssignalkarakteristika (feltintensitet, varighed, frekvens, modulationer, reelle eller simulerede), der påvirker sandsynligheden for at påvise RF-EMF-inducerede genetiske skader og dens forbindelse til termiske eller ikke-termiske interaktioner
5. potentielle biologiske mekanismer, der kan forklare RF-induceret genotoksicitet.
Sekundære spørgsmålspunkter
Yderligere spørgsmål har til formål at forklare årsagerne til modstridende resultater og uoverensstemmelser i tidligere studier:
6. Hvad afslører evidensen om risikoen for bias i studiedesign samt indflydelsen af finansieringskilder og forskertilknytninger på studieresultater?
7. Hvordan ændrer anvendelsen af strengere kvalitetskriterier evidensbalancen?
8. Er der tilstrækkelig homogenitet i dataene til at udføre en metaanalyse i et fremtidig systematisk review?
9. Har den eksisterende evidens huller, der kræver yderligere udforskning med fremtidsfokuseret forskning?
PECO-erklæring
Populationer: alle organismer og celletyper, mikroorganismer eller frit DNA, der anvendes i in vitro-, in vivo– og epidemiologiske studier.
Engagementer: Menneskeskabte radiofrekvenser (3 kHz til 300 GHz) fra virkelige trådløse sendere og signalgeneratorer (eller andre metoder, der bruges til at simulere virkelige enhedssignaler i laboratoriemiljøer).
Komparatorer: Sammenlign populationer udsat for lavere niveauer, falsk eksponering eller ingen eksponering (kontrol).
Resultater: DNA-skader som afbildet i Figur 1 samt relaterede potentielle biologiske mekanismer såsom spindelforstyrrelser, produktion af frie radikaler/oxidativt stress, varmechokproteinekspression og apoptose.
Metoder
Den protokol, der anvendes til at generere dette evidenskort og tilhørende datasyntese, følger udvidelsen Preferred Reporting Items for Systematic Reviews and Meta-Analyses for Scoping Reviews (PRISMA-ScR) (32) og Joanna Briggs Institute (JBI) retningslinjer for afgrænsningsreview (33). En fuldstændig beskrivelse af den anvendte metode og protokol findes i Supplerende datablad 1.
Analyse af undersøgelsens kvalitetsvurdering
Der blev foretaget en kvalitetsvurdering af studier med specifikke kvalitetskriterier. Vurderingen vedtog og udvidede de anbefalede kvalitetsegenskaber, der blev anvendt af Vijayalaxmi og Prihoda (30). De specifikke kvalitetsegenskaber, der anvendes til udvælgelse af studier af “højere kvalitet”, der er omfattet; bølgeform specificeret, eksponeringsvarighed beskrevet, detaljer om eksponeringsintensitet, signalfrekvens kvalificeret, dosimetri beregnet eller målt, blænding/kodning brugt, simuleret kontrol og statistiske metoder beskrevet. En positiv kontrol blev ikke vurderet til at være kritisk vigtig for kvalitetsbestemmelse, hvilket er en afvigelse fra Vijayalaxmi og Prihoda kvalitetsvurderingsprotokol. Mens positive kontroller er ønskelige for at evaluere et assays følsomhed ved at sammenligne effekter med et kendt genotoksisk middel, er de ikke afgørende for at bestemme årsag og effekt ved hjælp af eksperimentel logik (34). Studier, der inkorporerede alle disse kriterier, blev klassificeret som af højere kvalitet (n = 130). Yderligere oplysninger om kvalitetsvurderingsprocedurer findes i Supplerende datablad 1.
Fund i reviews
Kortlægning og præsentation af studier
Den vigtigste evidens for genotoksicitet, potentielle biologiske mekanismer og finansieringspåvirkninger opsummeres i et evidenskort ved hjælp af tabeller, grafer, et flowdiagram, varmekort og netværksdiagrammer. Den systematiske kortdatabase, der indeholder de udvalgte artikler, herunder bibliografiske oplysninger og udtrukne data, er opsummeret nedenfor, med alle detaljer i Supplerende datablad 4.
I alle grafer og tabeller viser hovedanalysen andelen af statistisk signifikante skadeseffekter (vist med orange) vs. andelen af ingen signifikant skade (vist i gråt). Alle tendenser i studier blev klassificeret som ikke-signifikante effekter, og studier, der viste tendenser eller beskyttende effekter, blev grupperet under kategorien “ingen effekt”, hvilket indikerer ingen signifikant DNA-skade.
Resultater fra søgning og screening (PRISMA)
PRISMA 2020-flowdiagrammet (Figur 2) giver et overblik over søge- og screeningsprocessen. En indledende søgning blev udført den 12. april 2023, begyndende med EMF-Portal-databasen. I alt 3.430 kandidatposter blev identificeret fra alle litteraturdatabaser tilsammen. Efter at have fjernet dubletter (n = 1674) og ikke-relevante eller ufuldstændige artikler (n = 1.226), var der i alt 530 artikler tilbage. Specifikke detaljer om artikelundtagelser findes i Supplerende datablad 4 (regnearksfane mærket “Gennemse”, kolonne B:C).
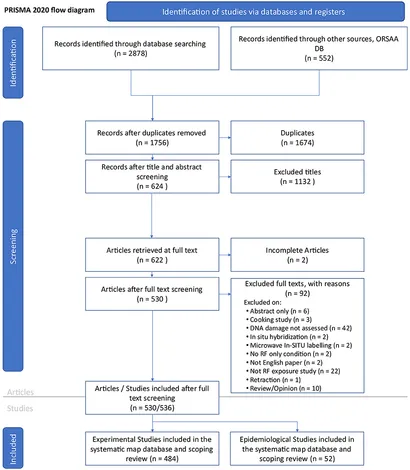
Fem artikler rapporterede om både in vitro- og in vivo-studier, og en artikel dækkede et epidemiologisk studie og in vitro-studie (hvilket bringer det samlede antal studier, der skal reviewess, n = 536). Af disse rapporterede nitten studier (6 % af in vitro og 1 % af in vivo) kun “mulige” DNA-skader og blev derfor udelukket fra den primære balance-of-evidens-kortlægningsproces (efterlader n = 517 studier til review).
Geografisk placering og publikationshistorie
Forskning i RF-EMF-genotoksicitet er blevet udført globalt, hvor USA, Kina, Indien, Italien, Japan og Tyrkiet er fremstået som de førende lande på dette område (Figur 3A). En mere detaljeret opdeling efter land og dokumentation findes i Supplerende tabel 2.
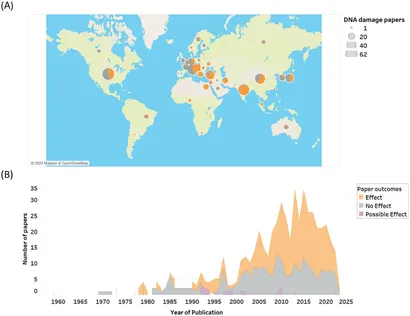
Over 500 studier af RF-EMF-induceret genotoksicitet og i mindre grad mutationer er blevet offentliggjort siden 1959 (Figur 3B). Antallet af forskningspublikationer forblev relativt stabilt fra slutningen af 1970’erne til 2000 og steg derefter i løbet af de næste 15 år, hvilket faldt sammen med den stigende anvendelse af trådløse enheder, udviklingen af understøttende infrastruktur og stigende folkesundhedsproblemer.
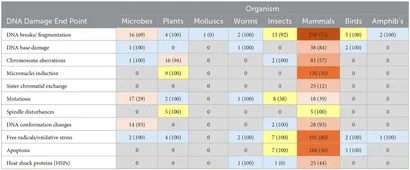
Organisme og celletyper
Genotoksicitetsforskning har primært fokuseret på pattedyr, hvor ikke-pattedyrsorganismer får mindre opmærksomhed. Bortset fra mikrober, planter og insekter (som har over 10 studier for nogle DNA-skadeslutpunkter), er mange organismer enten underundersøgt eller ikke undersøgt. Pattedyrsstudier (448 tilsammen) har primært fokuseret på forskning på mennesker (227 studier), hvor størstedelen er in vitro-studier og epidemiologiske studier, med færre in vivo-studier. Rotter er de næstmest udbredte (102 studier), derefter mus (78 studier) og andre pattedyr (41 studier), herunder et lille antal studier med kvæg, hunde, kattedyr, hamstere og kaniner (se Supplerende tabel 3). Forskningen dækker også en bred vifte af mikrobestudier (n = 33), herunder bakterie-, gær- og bakteriofagstudier (alle in vitro), som har vist blandede resultater (se Supplerende tabel 4).
Humane studier viste en næsten jævn fordeling (51 vs. 49 %) i evidensbalancen, hvor in vivo og epidemiologisk forskning konsekvent rapporterede statistisk signifikante DNA-skader sammenlignet med flere nulresultater for in vitro-studier. Rotter (75%) og andre pattedyr (73%) viste lignende tendenser. Mens resultaterne af musestudier generelt er på linje med resultaterne af humane studier, rapporterede en lavere andel af in vitro– og in vivo-musestudier DNA-skader. I modsætning hertil rapporterede alle studier af planter, orme, fugle og padder statistisk signifikante DNA-skader. Tilsvarende identificerede 71 % af insektunderstudier statistisk signifikante DNA-skadeeffekter.
Organisme vs. DNA-skadetype og -mekanisme
Cirka 80-100 % af de ikke-pattedyrsstudier viste statistisk signifikante effekter, især DNA-baseskader og oxidativt stress (Tabel 1). Spindelforstyrrelser, en potentiel mekanisme for DNA-skader, blev fundet i 100 % af studierne for både pattedyr og planter. Selvom de er mindre i antal (så konklusionerne er mindre sikre), har studierne med planter, insekter, orme og fugle høje andele af resultater, der viser RF-induceret DNA-skade og oxidativt stress. På grund af det begrænsede antal studier af padder, orme og snegle er det ikke muligt at drage solide konklusioner.
Skader efter celletype
In vitro-studier fokuserer primært på cellulær skade på tværs af forskellige organismer. På samme måde undersøger in vivo og visse epidemiologiske studier også specifikke celletyper. Når der observeres statistisk signifikant DNA-skade i > 50 % af studierne for bestemte celletyper, anses de for at være mere følsomme over for RF-EMF-eksponeringer.
Genotoksicitetsfund for forskellige celletyper afslørede varierende følsomhed over for RF-eksponering (se Tabel 2). Kønsceller var meget følsomme med statistisk signifikante DNA-skadeeffekter i 80 % af 20 æggestokke, 80 % af 30 testikler og 74 % af 27 spermatozostudier. Insektlarver (75 % af 4 studier) og embryoner (55 % af 20 studier) viste også forskellige niveauer af følsomhed. Normale hjerneceller viste sig at være meget følsomme (76 % af 54 studier), mens neoplastiske hjerneceller udviste lavere følsomhed (33 % af 12 studier). En moderat andel af lever- og lungestudier (56 % af 16 studier) fandt skader, såvel som studier af bukkale (orale) slimhindeceller (65 % af 23 studier). Det giver anledning til potentielle bekymringer på grund af ændringer i mobiltelefonteknologien, som flyttede hovedantennen fra toppen af telefonen til bunden, tættere på munden (36).
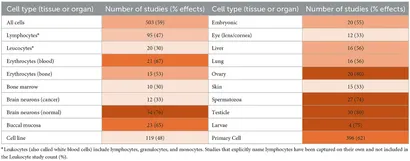
Blodceller viste større tolerance over for RF-induceret genotoksicitet, især lymfocytter (47% af 95 undersøgelser) og leukocytter (30% af 20 studier), som blev analyseret separat. Denne tolerance kan dog være begrænset, fordi længere eksponeringer, især i virkelige omgivelser (f.eks. epidemiologiske studier), viste statistisk signifikante DNA-skader på tværs af alle blodcelletyper. På den anden side fandt 67 % af 21 erytrocytstudier tegn på genotoksicitet. Nogle studier, der undersøger genotoksicitet på humant blod (37–39) viser også, at der er individuelle følsomheder og reaktioner på radiofrekvente eksponeringer, som ikke må forveksles med elektromagnetisk overfølsomhed (EHS). Nogle studier samlede deres resultater og udvaskede potentielt følsomme individuelle data (40–42).
Øjen- og hudvæv blev undersøgt sjældnere, hvor kun 33 % af henholdsvis 12 og 15 studier viste statistisk signifikante DNA-skadefund. Tilsvarende viste knoglemarv (30 % af 10 studier) mindre skade. Primære celler var mere tilbøjelige til at præsentere DNA-skader end cellelinjer. Disse resultater afslører variationen i cellefølsomhed.
Undersøgelsestype og fund af DNA-skade
Evidensfordeling efter undersøgelsestype (in vivo, in vitro, epidemiologisk)
Af de 517 inkluderede studier, der blev bekræftet at undersøge faktiske DNA-skader, var størstedelen in vitro-studier, der omfattede 53 % (n = 272) af alle studier. Til sammenligning var kun 37 % (n = 193) in vivo-studier, og mindre 10 % (n = 52) var observationsstudier (epidemiologiske). Der var fem artikler, der undersøgte både in vitro og in vivo resultater. Den samlede balance af evidens for DNA-skader var 59 % Effekter sammenlignet med 41 % Ingen effekter (Figur 4). Resultaterne for in vitro-studier blev vægtet en smule i forhold til ingen effekt (55 %), mens størstedelen af både in vivo-studierne (75 %) og de epidemiologiske studier (75 %) rapporterede statistisk signifikante DNA-skader. For studier af højere kvalitet skiftede balancen af evidens for den samlede DNA-skade til fordel for ingen effekter (52 %) vs. effekter (48 %).
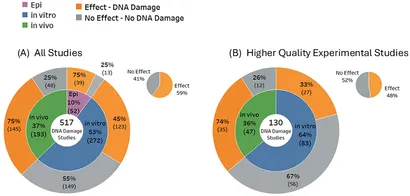
Disse resultater afslører betydelig variation i potentialet for, at RF-EMF kan inducere genetisk skade. In vitro-studier, som dominerer evidensgrundlaget, indikerer overvejende ingen signifikant DNA-skade. I modsætning hertil indikerer en identisk andel af in vivo-studier og epidemiologiske studier primært genotoksiske effekter. Disse modsatrettede resultater fra studier af levende organismer vs. isolerede celler fremhæver begrænsningerne ved in vitro-modeller til at replikere kompleksiteten af levende organismer, især dyr (44).
Dyr består af sammenkoblede systemer, herunder nervesystemet, det endokrine og immunsystemet, der alle kan påvirke cellulære funktioner og responser (45). Disse systemer formidler indviklede feedbackmekanismer, hormonel signalering og immunresponser (46). Desuden inkluderer in vivo-modeller fysiologiske processer og interaktioner såsom blodgennemstrømning, metabolisk aktivitet og interaktioner på vævsniveau, der kan påvirke RF-absorption og dens biologiske effekter (47, 48).
In vitro-systemer er imidlertid blottet for disse regulatoriske påvirkninger, interagerende systemer og processer, hvilket giver et begrænset og potentielt vildledende perspektiv på, hvordan celler reagerer på RF-eksponering i en levende organisme. Derudover kan det forenklede miljø for in vitro-studier føre til en undervurdering af RF-inducerede genotoksiske effekter. For fuldt ud at forstå den biologiske effekt af RF-eksponeringer skal in vitro-studier fortolkes forsigtigt og valideres gennem in vivo-forskning.
Epidemiologisk forskning afspejler på den anden side eksponeringsforhold i den virkelige verden og indikerer konsekvenserne af langvarig eksponering. Som forskningskortet viser (Figur 4), er epidemiologisk forskning mindre almindelig, potentielt på grund af kompleksitet, omkostninger og tidskrav. Evaluering af in vivo-data og epidemiologiske data resulterer tilsammen i et holistisk syn og modstandsdygtig bekræftelse. Derfor bør der lægges mere vægt på disse studietyper, når man vurderer sundhedsrisiciene ved RF-eksponering.
Typer af DNA-skader
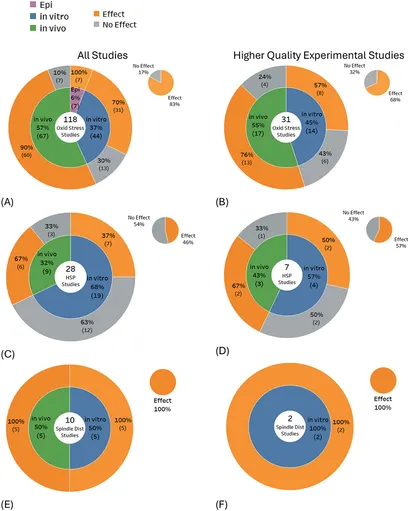
Statistisk signifikant DNA-skade var fremherskende i studier, der undersøgte DNA-brud (58 % af 283 studier), DNA-baseskader (86 % af 42 studier), kromosomafvigelser (63 % af 100 studier) og mikrokerneinduktion (54 % af 140 studier). DNA-baseskade blev fundet i de fleste studier for alle undersøgelsestyper: 92 % af 12 in vitro-studier, 80 % af 20 in vivo-studier og 100 % af 4 epidemiologiske studier (Figur 5C), og der blev fundet DNA-brud/-fragmenteringer i de fleste in vivo-studier (Figur 5A). Omvendt blev evidensbalancen vægtet mod ingen signifikante effekter for studier, der undersøgte mutationer (60 % af 45 studier) og søsterkromatidudveksling (88 % af 25 studier).
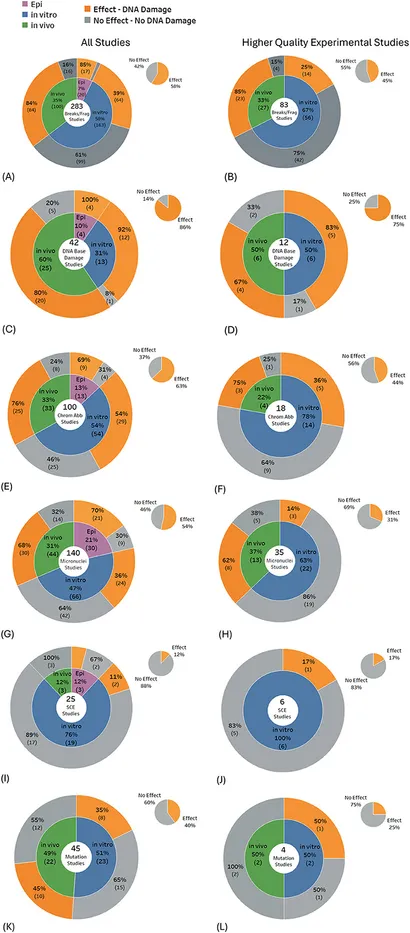
Når kun studierne af højere kvalitet blev udvalgt blandt ovenstående DNA-skadesubtyper, viste resultaterne typisk < 50 % af studierne, der rapporterede skader, bortset fra DNA-baseskader, hvor 75 % af de 12 studier af højere kvalitet gav statistisk signifikante resultater. Det største skift i evidensbalancen på tværs af de fleste DNA-skadesubtyper skete for in vitro-eksperimenter, hvor andelen af studier, der ikke viste nogen effekter, steg markant for studier af højere kvalitet. I modsætning hertil forblev evidensbalancen for in vivo-studier stort set uændret (se Figur 5A–L, grå områder i hvert cirkeldiagram).
Yderligere indikatorer for mulig DNA-skade
Nogle observerede biologiske ændringer tyder på DNA-skader, men uden nogen klar en-til-en overensstemmelse mellem forekomsten af disse biologiske ændringer og DNA-skader. Dokumentationen for disse mulige indikatorer blev ikke medtaget i den systematiske hovedkortlægning, der er beskrevet ovenfor, men præsenteres i stedet i Figur 6A–D for fuldstændighed. Studier, der undersøgte disse indikatorer, fandt overvejende DNA-konformationsændringer (93 % af 46 studier) og celleapoptose (60 % af 111 studier). Udvælgelse af kun studier af højere kvalitet havde minimal indvirkning på evidensbalancen for DNA-konformationsændring, dvs. resultaterne steg (100 % af otte studier), men for apoptosestudier faldt resultaterne (50 % af 26 studier). DNA-konformationsændringer viste en konsekvent højere andel af studier, der viste DNA-ændringseffekter for både in vitro og in vivo studier af højere kvalitet, mens for apoptose faldt statistisk signifikante fund for både in vitro og in vivo studier.
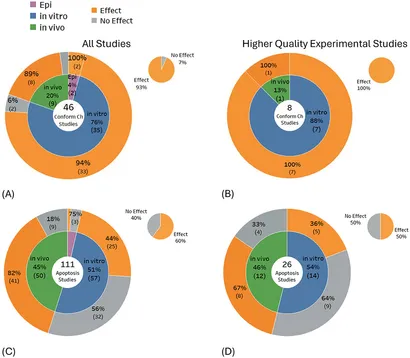
Synergistiske, additive og beskyttende virkninger
Additive, synergistiske, antagonistiske og potentiative effekter kan observeres og er afhængige af eksponeringssekvensen (før, efter eller samtidigt) og den oprindelige funktionelle tilstand af det eksponerede biologiske system [(49), s. 915].
Studier blev udført for at undersøge adaptive eller beskyttende celleresponser på radiofrekvenseksponering i forbindelse med kendte genotoksiske stoffer. Det blev kontrasteret med studier, der viste synergistiske effekter. Tilstedeværelsen af nogle midler (såsom gammastråler) i begge kategorier understreger den centrale indflydelse af timingen og varigheden af en RF-eksponering for at bestemme, om den cellulære respons sandsynligvis vil være beskyttende eller synergistisk (skadelig) (Supplerende tabel 11).
I nogle tilfælde indikerer de offentliggjorte data, at eksponering for RF-EMF kan give gavnlige beskyttende effekter mod andre genotoksiske stoffer, hvilket tyder på induktion af et adaptivt respons på både celle- og organismeniveau (50). Potentielle biologiske mekanismer, der ligger til grund for disse beskyttende effekter, omfatter aktivering af signalveje, opregulering af specifik genekspression og proteinsyntese (f.eks. HSP’er, antioxidantenzymer, cellereparation), øget aktivitet af DNA-reparationssystemet og en reduktion i frie radikalniveauer (51).
Selvom puljen af studier, der undersøgte kombinative og synergistiske effekter, var relativt lille, afslører den, hvordan RF-eksponeringer kan have potentielle terapeutiske fordele (kræftbehandlinger) eller kan øge skaden forårsaget af ioniserende stråling og kemiske stoffer, afhængigt af eksponeringsforholdene. Desværre tager de nuværende internationale RF-retningslinjer ikke hensyn til skade forårsaget af synergistiske interaktioner mellem RF og andre genotoksiske stoffer (52).
Karakteristika for eksponeringssignaler
Sammenhængen mellem eksponeringskarakteristika og genotoksicitetsfund er kortlagt i afsnittene nedenfor.
Eksponeringsfrekvens
Figur 7 opsummerer forskning og genotoksicitetsresultater efter frekvensbånd. De mest undersøgte bånd inkluderer 900-999, 1800-1899, 1900-1999 og 2400-2499 MHz, som stemmer overens med almindelige frekvenser, der bruges af mobiltelefoner, basestationer, mikrobølgeovne og Wi-Fi. Balancen af evidens favoriserer DNA-skadeeffekter for 900, 1800 og 2450 MHz frekvensbåndene. Resultaterne for 1900 MHz er dog tvetydige.
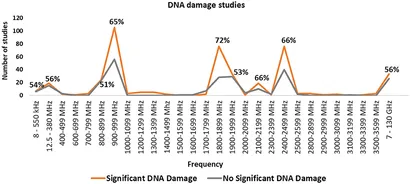
Yderligere studier, der dækker MHz- og GHz-områder, svarende til FM-udsendelser, magnetisk resonansbilleddannelse (MRI), radar og satellitkommunikation, er anført i Supplerende figur 3. Mange trådløse kommunikationsfrekvenser, især dem, der bruges af 5G new radio, er dog stadig underundersøgte. Som følge heraf anvendes nye frekvenser og modulationsordninger uden tilstrækkelig test for genotoksicitet eller bredere sundhedsmæssige konsekvenser.
Eksponeringens varighed
Dataene blev undersøgt for effekterne af eksponeringens varighed på studieresultaterne. Andelen af stuer, der viste statistisk signifikante DNA-skadeeffekter, blev tegnet over en bred vifte af eksponeringsvarigheder (1 minut til 1 år) for alle eksponeringsintervaller, der indeholdt fem eller flere studier.
Figur 8A tyder på et groft U-formet dosisresponsforhold, hvor over halvdelen af studierne i hver tidsgruppe viste effekter for både korte (mindre end en halv time) og længere eksponeringer (mere end 2 dage), mens mindre end halvdelen af studierne med eksponeringsvarigheder mellem en halv time og 2 dage viste effekter.
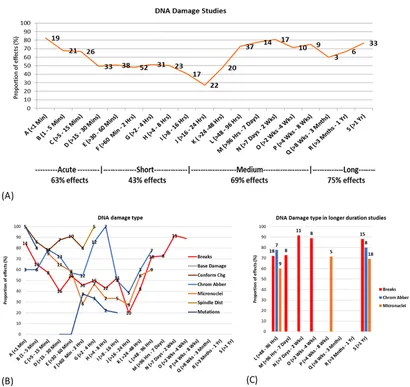
Både in vitro– og in vivo-studier viste den mindste mængde DNA-skader i studier i tidsintervallet 16-24 timer, hvor < 30 % af studierne viste DNA-skader. Mere end 50 % af in vitro– og in vivo-studierne viste statistisk signifikant DNA-skade ved eksponeringer < 15 minutter og >96 timer. Intervallet for eksponeringstid, hvor >50 % in vivo-studier viste skader, er også meget bredere end for in vitro-studier (se Supplerende tal 17a, 18a).
Resultaterne af eksponeringsvarigheden blev derefter undersøgt i henhold til specifik DNA-skadetype. Figur 8B illustrerer procentdelen af studier, der rapporterer effekter, der hver især tyder på en bi- eller trifasisk responskurve for de fleste DNA-skadetyper. Dette giver evidens for potentielle cellulære adaptive responser, især i tidsintervallerne 1-2 timer og 16-24 timer, hvor en større andel af studierne ikke rapporterede nogen effekter. Figur 8C fokuserer på de eksisterende studier med længerevarende eksponeringer (2 dage eller mere), der viser høje andele af statistisk signifikante effekter for DNA-brud, kromosomafvigelser og induktion af mikrokerner.
Kategorier af eksponeringsvarighed
For at forenkle analysen blev eksponeringsvarighederne kategoriseret i fire grupper: akut (167 studier), kortvarig (243 studier), mellemlang sigt (108 studier) og langsigtet (70 studier) (se Figur 9C). De fleste eksperimenter (70 %) fokuserede på kortvarige og akutte eksponeringer, hvor mellem- og langvarige eksponeringer var mindre undersøgt (30 %). In vitro-studier fokuserede primært på akutte og kortvarige eksponeringer, mens in vivo-studier omfattede alle eksponeringsvarigheder, og epidemiologiske studier koncentrerede sig om langtidseksponeringer. DNA-skader blev rapporteret i 63 % af akutte, 43 % af kortsigtede, 69 % af mellemlange og 76 % af langtidsstudier (se Supplerende tabel 12 for yderligere oplysninger). Disse eksponeringsvarighedskategorier blev brugt til at analysere varighed i kombination med andre faktorer, som beskrevet nedenfor.
Eksponeringsintensitet og varighed
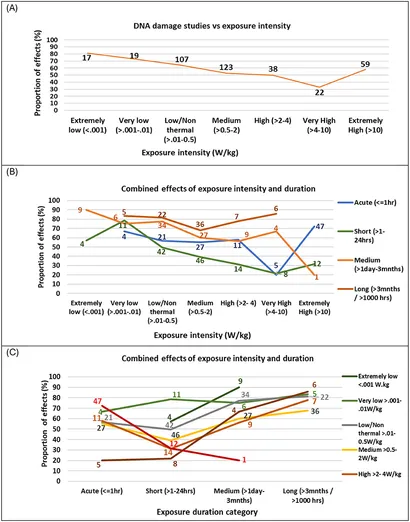
Dataene blev undersøgt for effekterne af eksponeringsintensitet på studieresultater ved at plotte andelen af studier, der viste effekter for en række eksponeringsintensitetsvinduer (ekstremt lave til ekstremt høje). Figur 9A illustrerer, at styrken af evidens for DNA-skader modereres af eksponeringsintensiteten på en ikke-lineær måde. De fleste studier har fokuseret på nærfeltsniveauer med lav til medium intensitet (>0,01-2,0 W/kg), typisk for mobiltelefonbrug, hvor halvdelen eller flere af disse studier fandt DNA-skadeeffekter. Færre studier har undersøgt højere eksponeringsintensiteter (f.eks. erhvervsmæssige indstillinger) eller ekstremt lave intensiteter, som er relevante for fjernfeltskilder såsom basestationer, smarte målere, radar og Wi-Fi-routere.
Studier, der undersøgte ekstremt lave eksponeringsniveauer (< 0,001 W/kg), rapporterede den højeste andel af genotoksiske effekter (81 % af 21 studier). Desuden faldt andelen af statistisk signifikante DNA-skadefund i takt med, at intensiteten steg. Ved ekstremt høje intensiteter (>10 W/kg), som overskrider ICNIRP-grænserne, steg andelen af statistisk signifikante DNA-skadeeffekter igen (58 % af 59 studier) (se Supplerende tabel 13). Disse resultater tyder på, at DNA-skader ikke følger et lineært dosis-respons-mønster, og tyder også på, at ikke-termiske mekanismer sandsynligvis vil spille en væsentlig rolle i RF-induceret DNA-skade. Efterfølgende blev eksponeringsintensiteten yderligere undersøgt ved at kategorisere eksponering i forhold til ICNIRP’s erhvervsmæssige grænser. Overraskende nok viste studier på eller under ICNIRP-grænserne en lidt større andel af statistisk signifikante genotoksiske effekter (58 % af 439 studier) end dem over grænseværdien (54 % af 92 studier) (se Supplerende tabel 13).
Eksponeringsintensitet og varighed blev yderligere undersøgt for at bestemme deres kombinerede effekt på genotoksicitetsfund. Figur 9B viser fordelingen af Figur 9A i kategorier af eksponeringsvarighed, hvis der er fem eller flere studier, der undersøger kombinationen af eksponering og varighed.
Andelen af studier, der viste effekter, varierede efter eksponeringens varighed og intensitet efter et mønster, der ligner Figur 9A. For studier med akutte (< 1 time) og korte (1-24 timer) eksponeringsvarighed viste en høj andel effekter ved ekstremt lave (< 0,001 W/kg) og meget lave (0,001-0,01 W/kg) intensiteter, men denne andel faldt, efterhånden som intensiteten steg. Disse studier nåede et minimum ved meget høje intensiteter (4-10 W/kg), med færre studier, der viste effekter, efterfulgt af en øget andel af effekter ved ekstremt høje intensiteter (>10 W/kg). For mellemlange studier (1 dag-3 måneder) nåede andelen af studier, der viste effekter, et indledende lavpunkt ved høj intensitet (2-4 W/kg), men forblev over 50 %. Ved ekstremt høje intensiteter (>10 W/kg) viste kun en ud af fem mellemlange studier effekter. I modsætning hertil udviste langtidsstudie (>3 måneder) en anden tendens: andelen, der viste effekter, var lavest ved medium intensitet (0,5-2 W/kg), men steg ved højere intensiteter. Især viste langvarige studier konsekvent effekter i over 65 % af tilfældene på tværs af alle intensitetsniveauer.
Alt i alt varierede effektmønsteret i et U-formet mønster på tværs af studievarigheden, med den højeste andel af effekter for lange (>3 måneder) og derefter mellemlange (< 1 dag-3 måneder) varighed, færre effekter for kortvarige studier (1-24 timer) og en større andel af effekter igen for akutte (< 1 time) varigheder. Figur 9C viser dette forhold tydeligere, idet det viser, at andelen af studier, der viste effekter, for alle eksponeringsintensiteter var mindst for korte eksponeringsvarigheder (1-24 timer). Derfor opstod to U-formede dosis-responsmønstre, et for intensitet og et for varighed, der interagerede med hinanden for at give et overordnet ikke-lineært og ikke-monotont dosisresponsmønster. Dette komplekse samspil bekræfter tidligere indikationer fra Lai og Levitt 2022 (43) af ikke-lineære responsmønstre for både intensitet og varighed. Det afslører, at intensitet alene ikke er den eneste vigtige faktor, der bestemmer resultaterne, og at eksponeringens varighed er en afgørende, modererende faktor. Desuden er lineære modeller ikke egnede til at beskrive resultater på dette område.
Disse resultater udfordrer de alt for forenklede konklusioner om ingen effekt fra nogle tidligere studier, som måske ikke har taget højde for det komplekse samspil mellem eksponeringsintensitet og varighed. De giver også retning for fremtidig forskning for yderligere at udforske disse ikke-lineære relationer.
Modulationer og simulerede signaler
Studier blev undersøgt for effekterne af forskellige telekommunikationssignalmodulationer (se Supplerende tabel 15). En større andel af studier, der brugte rigtige mobiltelefonsignaler, fandt DNA-skader, bortset fra GSM-Basic og CDMA kommunikationsprotokoller, hvor det lille antal studier begrænser evnen til at drage pålidelige konklusioner. Studier, der brugte simulerede signaler, var mindre konsistente, med 50 % eller mindre, der viste tegn på genotoksicitet på tværs af de fleste modulationsprotokoller. En undtagelse, GSM-Talk, viste statistisk signifikante DNA-skader for 67 % af 15 studier med simulerede signaler samt 91 % af 11 studier med et reelt signal. UMTS- og Wi-Fi-signaler udviste stærk genotoksicitetsevidens fra reelle signaler (88 % og 75 % af henholdsvis 8 og 12 studier), men mindre evidens for effekter (17 % og 33 % af henholdsvis 29 og 5 studier) for simulerede signaler. Samlet set viser disse resultater, at signalmodulation kan påvirke studieresultater og fremhæve de potentielle begrænsninger ved at bruge simulerede signaler til at evaluere genotoksiske risici.
Pulserende vs. kontinuerlige signaler
Pulserende og kontinuerlige bølger viste begge en større andel af DNA-skader for in vivo og epidemiologiske studier og en lavere andel for in vitro-studier. Kontinuerlige bølger producerede en større andel af DNA-skader end pulserende bølger for in vitro-eksperimenter (men stadig < 50 % af 116 studier). Når alle studier kombineres, har både pulserende og kontinuerlige bølgestudier en tendens til større DNA-skader. Flere eksponeringer, som involverer separate gentagne eksponeringer på en enkelt dag eller over flere dage, viste sig at give flere positive resultater end en enkelt eksponering.
Sammenligninger af eksponeringsmønstre, dvs. kontinuerlige, intermitterende (f.eks. 5 minutter tændt, 10 minutter slukket) og variable (dynamisk skiftende intensitet, såsom under brug af mobiltelefoner) afslører en progressiv stigning i andelen af DNA-skadefund: 55 % af 369 studier for kontinuerlige, 62 % af 60 studier for intermitterende og 76 % af 96 studier for variable eksponeringer.
Yderligere effekter af signalkarakteristika på resultaterne præsenteres i Tillægstabel 15–17. På tværs af alle eksponeringskarakteristika rapporterer in vivo-studier og epidemiologiske studier konsekvent en højere andel af DNA-skader end in vitro-studier.
Potentielle biologiske mekanismer
Mekanismer for DNA-skader
En gennemgang af studier, der undersøger potentielle biologiske mekanismer (Figur 10A–F) viste, at produktion af frie radikaler eller oxidativt stress var det hyppigst undersøgte endepunkt, og de fleste studier viste effekter (83 % af 118 studier). Varmechokproteiner blev fundet udtrykt i knap halvdelen af studierne (46 % af 28 studier), og spindelforstyrrelser blev fundet i alle relevante studier (100 % af 10 studier). For studier af højere kvalitet forblev evidensbalancen vægtet mod effekter for produktion af frie radikaler/oxidativt stress (68 % af 31 studier) og vippede mod effekter for varmechokproteinekspression (57 % af syv studier) og forblev uændret for spindelforstyrrelser (100 % af tostudier).
Mønstre i eksponeringskarakteristika
Samlet set afslører resultaterne, at forholdet mellem eksponeringsintensitet og DNA-skader varierer ikke-lineært med eksponeringsvarigheden. Evidensen er tungt vægtet af kortsigtede, in vitro-studier, hvor adaptive responser ser ud til at forekomme. Signaltype, frekvens og eksponeringsmønstre viste signifikant indflydelse på studieresultaterne. Almindeligt anvendte bånd som 900-999, 1800-1899 og 2400-2499 MHz, der bruges af mobiltelefoner, smarte enheder og Wi-Fi, viste større andele af effekter. 5G-bånd er på den anden side underundersøgt. Pulserende bølger fremkaldte en større andel af effekter for in vivo– og epidemiologiske studier. Enheder i den virkelige verden viste konsekvent en større andel af effekter end signalgeneratorer.
Alt i alt er de eksponeringskarakteristika, der er mere tilbøjelige til at fremkalde DNA-skadeeffekter, dem, der beskriver autentiske eksponeringsforhold. Disse resultater understreger vigtigheden af fremtidige studier for at inkorporere forskellige frekvenser, realistiske eksponeringsmønstre og nye teknologier.
Mekanismer og eksponeringsvarighed
Andelen af studier, der viste effekter på potentielle DNA-skademekanismer, blev sammenlignet for forskellige eksponeringstidsintervaller. Kun de tidsintervaller med fem eller flere studier (for effekter eller ingen effekter kombineret) blev analyseret.
Når mekanismer blev undersøgt på tværs af eksponeringstidsintervaller, blev et bi- eller trifasisk responsmønster foreslået (se Figur 11A), hvor frie radikaler/oxidativt stress var dominerende i studier, hvor der blev anvendt 30 minutter til 2 timers eksponering, og hvor frie radikaler/oxidativt stress og apoptose var dominerende i studier, hvor eksponeringsvarigheder var længere end 2 dage. Disse resultater følger de samme mønstre af effekter for DNA-skadetyper vist i Figur 11 (Se også Supplerende tal 21–27 for flere detaljer). De mulige sammenhænge mellem de underliggende mekanismer for DNA-skade og typen af DNA-skade blev efterfølgende undersøgt ved at overlejre andelen af statistisk signifikant evidens for hver på tværs af de forskellige eksponeringsvarigheder.
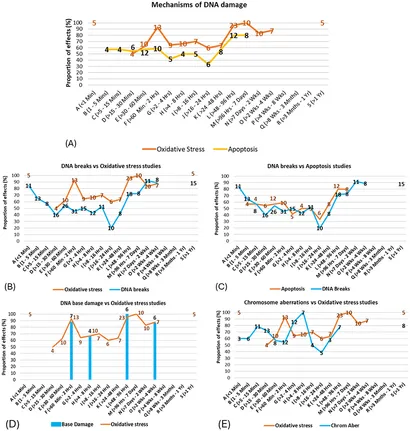
For eksempel viser studier af frie radikaler/oxidativt stress og DNA-brud lignende mønstre af resultater på tværs af tidsintervallerne (Figur 11B), hvilket tyder på, at produktion af frie radikaler/oxidativt stress kan være årsagsrelateret til DNA-brud. Tilsvarende Figur 11 antyder stærke korrespondancer mellem (C) apoptose og DNA-brud/fragmenteringer og korrespondancer mellem (D) frie radikaler produktion/oxidativ stress og DNA-baseskade.
Potentielle forbindelser blev også observeret mellem (E) produktion af frie radikaler/oxidativ stress og kromosomafvigelser, med en tidsforsinkelse mellem førstnævnte og sidstnævnte; Der er dog brug for mere evidens for at styrke denne formodning. Der blev også observeret mulige yderligere sammenhænge mellem spindelforstyrrelser og kromosomafvigelser, kromosomafvigelser og mikrokerneinduktion og mellem produktion af frie radikaler/oxidativ stress og både kromosomafvigelser og mikrokerneinduktion (se Supplerende tal 23–26 for alle detaljer). Overensstemmelserne mellem mønstrene af evidens for mekanismer og DNA-skadeeffekter i specifikke tidsintervaller understøtter resultatet fra Figur 9 ovenfor, samt eksisterende teori, der tyder på, at effekter fra EMF-RF-eksponeringer er stærkere i visse tidsvinduer (53, 54).
Produktion af frie radikaler som en potentiel mekanisme
Figur 11B og Supplerende figur 30 illustrerer, at der er en stærk overensstemmelse mellem at observere frie radikaler/oxidativt stress og finde DNA-skader. Evidens for oxidativ DNA-baseskade viste den højeste andel af statistisk signifikante fund blandt alle undersøgte former for DNA-skade (Figur 5C). To kritiske markører for oxidativt stress og carcinogenese (55), var 8-oxo-7,8-dihydro-2′-deoxyguanosin (8-oxo-dG) og 8-hydroxy-2′-deoxyguanosin (8-OHdG) de hyppigst målte biomarkører for oxidativ DNA-skade. Begge disse oxidative DNA-skadebiomarkører ses som risikofaktorer for mange sygdomme, herunder neurodegenerative lidelser (56) og kræft (57). Manglende øjeblikkelig fjernelse af 8-oxo-dG kan resultere i en basetransversionspunktmutation, hvor G:C omdannes til T:A under DNA-replikation (56).
Reaktive iltarter (ROS) er frie radikaler, der almindeligvis observeres som et endepunkt i adskillige RF-eksponeringsstudier (58), hvor de ofte er forbundet med oxidativt stress. Forhøjede markører for oxidativt stress er blevet fundet hos personer med neurologiske lidelser såsom Alzheimers sygdom og Parkinsons sygdom samt diabetes, hjerte-kar-sygdomme og kræft (59). Små stigninger i ROS er også blevet observeret for at give terapeutiske effekter; Disse effekter er dog kun tydelige inden for snævre eksponeringsintensitetsvinduer (60) og varighed.
Der er identificeret flere kendte og potentielle veje, hvormed RF-EMF-eksponering kan føre til en ubalance i frie radikaler i celler (se også Figur 12):
• Mitokondriel dysfunktion (61, 62);
• RF-forbedrede Haber-Weiss- og Fenton-reaktioner (H2O2, OH−, Fe2+ og Cu+ ioner) (63–65);
• Mikrobølgeinteraktioner med vandmolekyler for at danne H2O2 (66, 67);
• Ændret antioxidant genekspression (68).
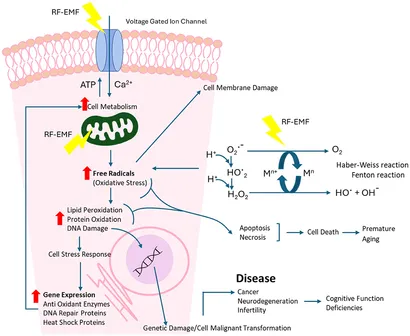
Reaktive iltarter er også involveret i aktiveringen af cellulære signalveje, herunder reguleringen af de vigtigste veje for apoptose, især den mitokondriemedierede vej (69) (se Figur 12). Øgede niveauer af reaktive iltarter er blevet forbundet med DNA-skader og forbundet med miljøstress, hvor menneskeskabte radiofrekvenseksponeringer er en bidragyder (70).
Bifasiske forhold på grund af cellulære adaptive responser
Celler anvender forskellige beskyttelsesmekanismer, når de står over for cellulær stress, herunder opregulering af DNA-reparationsgener, varmechokproteiner og enzymer, der afbøder oxidativt stress (71, 72). Genekspression er en sekventiel proces, der kræver tid, startende med signalvejsaktivering og transkriptionsfaktorbinding, efterfulgt af mRNA-behandling og proteinsyntese (73). Den tidsafhængige aktivering af cellulære stressresponser kan forklare den U-formede eller bifasiske dosisrespons på RF-EMF-eksponering, der observeres over stigende eksponeringsvarigheder (Figur 11). Et nyligt studie, der undersøger produktion af frie radikaler, har bekræftet en bifasisk cellulær respons på RF-EMF-eksponering (74).
Akutte (meget korte) eksponeringer for ekstremt høje RF-EMF-intensiteter resulterer ofte i statistisk signifikante DNA-skader, muligvis på grund af reparationsmekanismernes manglende evne til at reagere hurtigt. I modsætning hertil kan længere eksponering for høje intensiteter føre til aktivering af cellulært forsvar for at reparere skader og forsøge at afbøde skade. Desuden kan langvarig eller kumulativ eksponering føre til akkumuleret DNA-skade, genomisk ustabilitet og cellulær dysfunktion (75). Afhængigt af skadens omfang kan celler aktivere autofagi som en beskyttelsesmekanisme til at fjerne beskadigede komponenter og opretholde cellulær homeostase (76, 77). Selvom det kan være effektivt på kort sigt, kan celler, hvis skaden fortsætter eller overvælder reparationsmekanismer, gennemgå apoptose for at fjerne alvorligt kompromitterede celler. Men hvis denne overlevelsesproces mislykkes, er der risiko for ondartet transformation (75).
RF-eksponeringer i den virkelige verden er typisk kroniske og variable, hvilket giver anledning til bekymring for kumulative effekter (78). Mens reparationsmekanismer ser ud til at løse meget af skaden på kort sigt, kan langvarige eller gentagne eksponeringer overvælde dette forsvar, hvilket fører til varige genomiske ændringer. Da en stor del af offentlighedens eksponeringer nu finder sted fra vugge til grav og ofte er uden samtykke, er der behov for at adressere de potentielle langsigtede sundhedsrisici ved vedvarende RF-eksponering.
DNA-konformationelle ændringer
DNA-konformationsændringer er en potentiel markør for DNA-skader. Forskellige metoder er blevet anvendt i studier til at identificere ændringer i DNA-konformation, herunder Raman-spektroskopi, elektronmikroskopiobservationer, cirkulær dikroisme og dynamiske lysspredningsteknikker samt UV-vis spektroskopi. Den mest almindelige anvendte metode brugte imidlertid Anomalous Viscosity Time Dependency (AVTD), som giver indsigt i strukturel dynamik og molekylære interaktioner inden for biologiske systemer. Denne metode viser især, hvordan DNA-proteinkomplekser (f.eks. kromatin) påvirker DNA’ets fysiske egenskaber, såsom viskositet i en opløsning over tid (79).
Kromatin er et DNA-proteinkompleks, der findes i kernen af eukaryote celler. Strukturen af kromatin kan gennemgå dynamiske ændringer som reaktion på forskellige cellulære processer, herunder DNA-reparation, transkription og replikation. Proteiner såsom histoner og ikke-histonkromatin-associerede proteiner spiller afgørende roller i organisering og regulering af kromatinstruktur (80).
Resultaterne viste, at RF-eksponeringer kan forårsage DNA-konformationsændringer med en høj grad af konfidensitet (93 % af studierne), (se Supplerende tabel 9). Selvom konsekvenserne af en sådan ændring ikke er fuldt ud forstået, er et muligt scenarie den situation, hvor DNA-skader bliver utilgængelige for reparationsproteiner, eller reparationsfunktionen hæmmes af ændringer i reparationsenzymer (81).
Mens AVTD og lignende metoder giver værdifuld indsigt i DNA-konformationsændringer, kan de ikke let bestemme den underliggende årsag, eller om den relaterer til DNA-skader. Integration af AVTD med andre eksperimentelle tilgange, såsom et kometassay, kan hjælpe med at belyse, om de observerede ændringer er direkte relateret til DNA-skader eller resultatet af normale cellulære reguleringsprocesser.
Forholdet mellem kromosomafvigelser, mikrokerner og spindelforstyrrelser
Mikrokerner, kromosomafvigelser og spindelforstyrrelser er alle indbyrdes forbundne indikatorer for genomisk ustabilitet og cellulær stress (82). Det mitotiske spindelapparat er ansvarlig for kromosomadskillelse under celledeling. Spindelforstyrrelser kan føre til forkert kromosomjustering og adskillelse, hvilket fører til forsinkede kromosomer eller fragmenter, der kan blive indkapslet og danne mikrokerner (82).
Der er tidligere identificeret en positiv korrelation mellem mikrokerner og særlige kromosomafvigelser i humane in vitro-studier, specifikt for acentriske fragmenter og dicentriske kromosomer (83). Desuden kan dannelse af mikrokerner induceres af kromosombrud eller hæmning af spindlerne under celledeling (84).
Et studie af graferne, der viser procentdelen af studier, der rapporterer mikrokerneinduktion, kromosomafvigelser og spindelforstyrrelser over eksponeringstiden, når de er overlejret på hinanden, viser et tæt forhold, der validerer udsagn fra tidligere forskere (se Supplerende tal 23, 25).
Proteiner til varmechok
Varmechokproteiner (HSP’er) er chaperoner, der beskytter cellemakromolekyler. De spiller en afgørende rolle i både reparation af DNA-skader og den cellulære reaktion på oxidativt stress (71, 72). Eksperimentel evidens tyder på, at RF-eksponering kan inducere HSP-ekspression, som beskytter celler, men paradoksalt nok kan også hjælpe kræftcellers overlevelse ved at hæmme apoptose. RF-eksponering har vist sig at opregulere HSP’er i humane endotelceller, hvilket potentielt bidrager til tumorprogression (85).
Blot 27 studier undersøgte forholdet mellem RF-EMF-eksponering og ekspressionen af varmechokproteiner, som omfattede HSP27, HSP70 og HSP90. Bortset fra ekstremt høje eksponeringsniveauer synes der at være en omvendt sammenhæng mellem eksponeringsintensitet og detekterede HSP-niveauer (se Supplerende tabel 20). Evidensen følger et lignende mønster som forholdet mellem eksponering og intensitet og DNA-skader (se Supplerende tal 20A, B og Supplerende tabel 13). Det begrænsede antal studier ved visse eksponeringsniveauer forhindrer imidlertid pålidelige konklusioner.
Apoptose: En konsekvens eller årsag til DNA-skader i RF-EMF-studier?
Apoptose er en normal, stramt reguleret proces, der bruges til at eliminere overflødige celler og beskadigede celler, herunder celler med DNA-skader, for at reducere risikoen for kræftfremkaldende (69). Den apoptotiske proces kan initieres af tre forskellige signalveje (86):
1. iboende (fremmes af mitokondrier)
2. ekstrinsisk (involverer dødsreceptorer på celleoverfladen) og
3. via det endoplasmatiske retikulum.
Caspase-enzymer er afgørende mediatorer af apoptose, der orkestrerer den ordnede demontering af cellulære komponenter under programmeret celledød. Deres aktivering er et kendetegn for apoptose (87), med flere studier i dette evidenskort, der anvender caspaseanalyser til påvisning af apoptotiske hændelser.
Analyse af resultaterne afslører en stærk overensstemmelse mellem DNA-skader og apoptose, som det fremgår af (Figur 11C), hvor adskillige studier rapporterer begge resultater efter RF-EMF-eksponering. Det tætte forhold rejser et vigtigt spørgsmål: Blev apoptose i disse studier udløst af overdreven DNA-skade fra RF-EMF, hvilket resulterede i kontrolleret celledød, eller var den påviste DNA-skade resultatet af apoptose? Begge scenarier er plausible. Apoptose er en cellulær mekanisme, der initieres, når akkumuleret skade, herunder DNA-skade, overstiger cellens evne til at reparere og komme sig (86). Den apoptotiske biokemiske proces kan omfatte DNA-spaltning ved at aktivere endogene endonukleaser (88), hvilket fører til DNA-fragmentering. Mange studier, der undersøgte apoptose, gav imidlertid ikke tilstrækkelige oplysninger til at verificere den primære initieringsmekanisme.
Studie kvalitet og resultater
En slående uoverensstemmelse blandt de gennemgåede studier var deres kvalitet, med adskillige udeladelser af kritiske detaljer såsom komplet studiemetodologi, bølgeegenskaber (f.eks. pulserende eller kontinuerlige bølger, anvendt modulering), falsk eksponering, blinding, tilstrækkelig dosimetri eller omfattende statistiske data.
Studieresultater blev undersøgt som en funktion af studiekvaliteten. Anvendelse af kvalitetskriterierne (se Metoder og Supplerende datablad 1) flyttede andelen af studier, der viste DNA-skader, fra 59 % af alle 517 studier til 48 % af 130 studier af høj kvalitet (se Figur 4).
De vigtigste kvalitetskriterier blev yderligere undersøgt separat for at se, hvordan de påvirkede resultaterne for de eksperimentelle studier. Resultaterne viste, at anvendelsen af hvert af kvalitetskriterierne individuelt reducerede andelen af studier, der viste effekter. Der var en lavere andel af studier, der viste effekter, når studierne var blindede (47 %) vs. ikke blindede (64 %), hvor tilstrækkelig dosimetri blev inkorporeret (56 %) vs. ikke inkorporeret (61 %), når der blev anvendt simulerede kontroller (50 %) i forhold til kun normale kontroller (70 %). Studier, der inkorporerede alle disse tre kriterier, viste en lavere andel af DNA-skader, 48 % (se Supplerende tabel 21).
Yderligere studier afslørede imidlertid, at dette skift mod færre statistisk signifikante resultater, når kvalitetskriterier blev anvendt, kun var sandt for in vitro-studier. Der var meget få ændringer i resultaterne, når kvalitetskriterier blev anvendt på in vivo-studier. I modsætning hertil steg andelen af studier, der viste effekter, en smule, når kurven blev specificeret, og andelen af studier, der viste ingen effekt, faldt en smule (se Supplerende figur 31).
De resterende kvalitetskriterier blev anvendt i for få studier til at muliggøre meningsfuld sammenligning, dvs. eksponeringsvarighed ikke beskrevet (n = 2), signalfrekvens ikke kvalificeret (n = 8). Forskning af højere kvalitet og lavere kvalitet inden for hver forskningsgruppe (finansieringstilhørsforhold) gav lignende resultater. Der opstod imidlertid et modsat mønster, når man sammenlignede balancen mellem evidensresultaterne mellem forskningen i egeninteresse og den uafhængige forskning (se Supplerende tabel 27).
Alle parametre og studiekvalitet
Ovenstående faktorer, der viser effekter på studieresultater, blev samlet undersøgt for eventuelle eksperimentelle parametre, der kan forklare den lavere andel af DNA-skadeeffekter, der findes i studier af højere kvalitet sammenlignet med alle studier. Det blev konstateret, at studier af højere kvalitet har:
• en lidt større andel af in vitro-studier, som har tendens til at vise nulresultater (se Tillægstabel 24a, b);
• en lavere andel af in vivo-studier, som typisk viser statistisk signifikante DNA-skader (se Tillægstabel 24a, b);
• en større andel af studier, der bruger simulerede signaler via en signalgenerator, hvilket yderligere skævvrider resultaterne i retning af ingen effekt (se Tillægstabel 24c, d);
• brugt flere cellelinjer, hvilket kan bidrage til mindre modtagelighed for RF-induceret DNA-skade (på grund af usikkerhed i cellearv, fænotype eller tidligere strålingseksponering) (se Tillægstabeller 24g, h);
• fokuserede mindre på akutte eksponeringer, hvor der ofte findes DNA-skader, og mere på kortvarige eksponeringer, hvor der kan være adaptive reaktioner, hvilket reducerer sikkerheden for resultaterne (se Tillægstabeller 23a–d);
• signifikante forskelle i antallet af studier, der er udført med mellemhøj og ekstremt høj intensitet, hvor studier af højere kvalitet fokuserede mere på studier med mellemhøj intensitet, hvor resultaterne for DNA-skader er tvetydige, og færre studier med ekstrem høj intensitet, hvor der er større sandsynlighed for at finde DNA-skader (se Tillægstabeller 22g, h, m, n);
• en større andel af forskningen i denne kategori, der potentielt er knyttet til særinteresser, samtidig med at der lægges større vægt på in vitro-studier (se Supplerende figur 53 og Supplerende tabel 24); og
• udelukkede epidemiologiske studier (på grund af dosimetrimangler), hvor der overvejende blev fundet DNA-skader.
Disse ekstra parametre, der er anført ovenfor, kan bidrage til den lavere evidensbalance for effekter, der er observeret i studier af højere kvalitet, lige så meget som de etablerede kvalitetskriterier (30).
Bekymringer om dosimetri
Trådløse enheder i den virkelige verden forventes at overholde de grænser for offentlig eksponering, der anbefales af ICNIRP (15). Beregning af dosimetri i studier, der bruger disse enheder, er imidlertid kompleks på grund af den variable karakter af de udsendte signaler (89). Mange studier viste sig at mangle tilstrækkelig information om dosimetri eller de metoder, der anvendes til beregning eller måling heraf.
Nogle forskere har kritiseret studier, der anvender enheder fra den virkelige verden for “dårlig dosimetri”, og brugt dette argument til at sætte spørgsmålstegn ved studiets kvalitet og nedtone resultaterne (30, 90). Der blev dog observeret minimale forskelle i evidensbalancen, når man sammenlignede såkaldte studier med “dårlig dosimetri” (som typisk repræsenterer eksponeringer under virkelige forhold) med dem med “tilstrækkelig dosimetri”. I betragtning af at disse umodificerede enheder i den virkelige verden fungerer inden for og ofte langt under ICNIRP’s offentlige eksponeringsgrænser, bliver kritikken af dårlig dosimetri mindre relevant, når man evaluerer gyldigheden af de nuværende offentlige sikkerhedsstandarder.
Risiko for bias og studieresultater
En af de mest kritiske udfordringer ved evaluering af det genotoksiske potentiale af RF-EMF er at identificere indflydelsen af finansieringskilde eller forfatteransættelsesforhold, som har potentiale til at skævvride forskningsresultater og hindre udviklingen af evidensbaserede politikker. Finansieringstilknytning(er) har stor indflydelse på de rapporterede resultater, som det fremgår af tidligere reviews (91–94). En omfattende analyse af potentielle bias, som undersøgte forfatternes fokusområder, finansieringskilder og tidsskrifter, der blev brugt til publicering, afslørede en stærk indflydelse på rapporterede resultater, som vist af datamønstrene nedenfor.
Undersøgelsesparametre vs. finansieringskilde
Diagrammerne præsenteret i Figur 13 Vis dokumentationsbalancen efter parameter, grupperet efter primær finansieringskilde. (Bemærk, at studier kan have flere finansieringskilder, herunder institutionelle, industrielle eller statslige. Derfor blev der anvendt et filter til at fokusere på én finansieringskilde for hver graf; Bemærk, at finansiering fra militære og telekommyndigheder udtrykkeligt blev udelukket fra statslig finansiering).
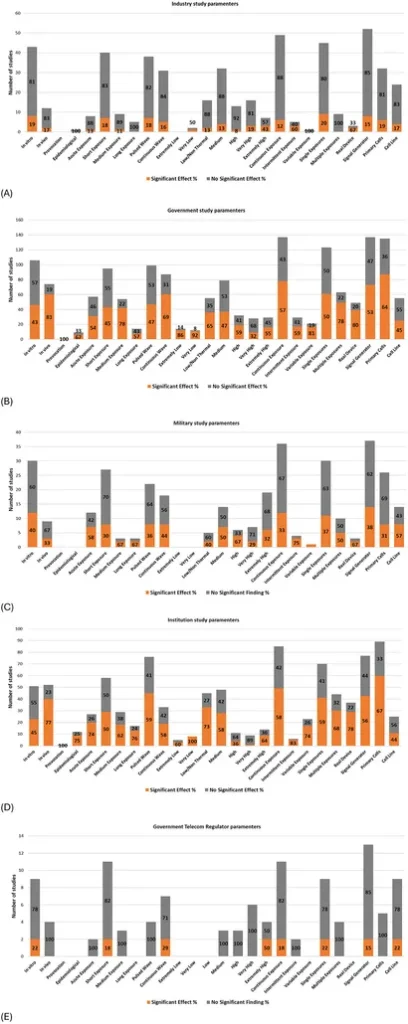
Orange: Effekt i % Grå: Ingen effekt i %.
Forskning finansieret af særinteresser (f.eks. industrien, statslige teletilsynsmyndigheder og i mindre grad militær – se afsnittet “Klassificering af potentielle særinteresser”; Supplerende datablad 1) viser en større andel af studier, der konkluderer “ingen DNA-skade” (som illustreret ved mængden af grå farve) sammenlignet med studier finansieret af regeringer, institutioner eller private/offentlige kilder (ikke vist). Især ligner studier finansieret af USAF industrifinansieret forskning mere end typiske militærfinansierede studier (se Supplerende datablad 5, Tillægstabel 25 og Supplerende figur 35 for alle detaljer).
Finansieringskilder synes at have fastlagt forskellige eksperimentelle parametre, der svarer til forskningsprioriteter og potentielle skævheder. Studier med potentielle interessekonflikter (COI), såsom dem, der er finansieret af industrien, militæret eller statslige teletilsynsmyndigheder, organisationer, der kollektivt har interesser i RF-teknologi, udviser følgende forskningstendenser:
1. Et stort antal in vitro-studier, begrænsede in vivo-studier og minimale epidemiologiske studier.
2. Overvægt af korte eksponeringsvarigheder (1-24 timer) med begrænsede studier af akutte, mellemlange eller langsigtede eksponeringer;
3. Afbalanceret studier af pulserende og kontinuerlige bølgeeksponeringer;
4. Undgåelse af eksponeringer med ekstremt lav intensitet med præference for eksponering af medium til høj intensitet
5. Vægt på kontinuerlig eksponering med minimal fokus på intermitterende eller variable eksponeringer;
6. Præference for enkelteksponeringer frem for multibestråling
7. Større afhængighed af signalgeneratorer sammenlignet med trådløse enheder i den virkelige verden; og
8. Afbalanceret brug af primære celler og cellelinjer.
Nogle af ovenstående undersøgelsesdesignbeslutninger er mere tilbøjelige til at producere nulresultater (som diskuteret i afsnittene ovenfor).
I modsætning hertil viser uafhængig forskning, dvs. forskning, der ikke er finansieret af industrien, teletilsynsmyndigheden eller militæret, en bredere dækning af studieparametre, der er kendetegnet ved:
1. Undersøgelsesdesign, der gør brug af hele spektret af undersøgelsesparametre, med næsten paritet mellem in vitro- og in vivo-studiemængden.
2. De fleste epidemiologiske studier blev udført som uafhængig forskning;
3. Kortsigtede eksponeringer er almindelige; dog er akutte, mellemlange og langsigtede eksponeringer bedre repræsenteret;
4. Lidt større fokus på pulserende bølgeeksponeringer sammenlignet med kontinuerlige bølger;
5. Omfattende brug af lav/ikke-termisk og mellemhøj eksponering med god dækning af andre intensiteter
6. Kontinuerlig eksponering er mere dominerende; dog er intermitterende og variable eksponeringer også blevet undersøgt;
7. Enkelteksponeringer er mere almindelige, men mindre dominerende sammenlignet med undersøgelser, der finansieres af kapitalinteresser.
8. Signalgeneratorer bruges oftere end trådløse enheder i den virkelige verden, men ubalancen er mindre udtalt end undersøgelser finansieret af særinteresser; og
9. Primære celler foretrækkes frem for cellelinjer.
Studier finansieret af industrien, telemyndighederne og USAF-finansierede studier har alle en lignende balance mellem evidensprofiler (Tillægstabel 25). I disse tilfælde viser de fleste studier (>80%) ingen signifikant DNA-skade. Der ses dog subtile forskelle mellem dem på det eksperimentelle niveau.
Effekten af finansiering på studieresultater forværres yderligere af manglen på gennemsigtighed og offentliggørelse, hvor 173 studier (~33 % af studierne) mangler en formel finansieringserklæring.
Potentielle interessekonflikter
Design og resultater af forskningsstudier, der modtog finansiering fra industrien, teletilsynsmyndigheder eller militæret, blev sammenlignet med forskningsstudier, der blev klassificeret som uafhængige. De to forskningsgrupper (potentiel egeninteresse vs. uafhængig) foretrak forskellige studietyper og viste modsatrettede proportioner af effekter i mange tilfælde (se Supplerende tabel 27). Forskning finansieret af eller tilknyttet industrien eller militæret omfattede primært in vitro-studier, hvor 30 % af 142 in vitro-studier og 26 % af 31 in vivo-studier rapporterede statistisk signifikante resultater. Kvalitetsfiltrering påvirkede ikke disse resultater signifikant (henholdsvis 25 % af 56 in vitro-studier og 25 % af 12 in vivo-studier).
I modsætning hertil blev 63 % af de 465 eksperimentelle studier klassificeret som uafhængig forskning med en blanding af 130 (44,5 %) in vitro-studier og 162 (55,5 %) in vivo-studier. Uafhængige in vitro-studier, uanset kvalitet, viste flere nulresultater end in vivo-studier. Samlet set havde studiekvaliteten ringe indflydelse på uafhængig forskning, hvor 74 % af alle uafhængige studier (n = 292) viste statistisk signifikante DNA-skader sammenlignet med 73 % af 62 studier af højere kvalitet.
Studier af højere kvalitet havde en tendens til at rapportere mere konservative resultater (mod ingen signifikante effekter), i overensstemmelse med tidligere RF-EMF-reviews af Wood et al. (95), Karipidis et al. (96), Vijayalaxmi og Prohida (30) og Simko et al. (97). Disse reviews vurderede dog ikke den kombinerede indflydelse af potentielle særinteresser (f.eks. finansiering eller forfattertilhørsforhold) og undersøgelseskvalitet på resultater. De betydelige forskelle mellem forskningsgruppens resultater (potentielle særinteresser vs. uafhængige) giver anledning til alvorlig bekymring om pålideligheden og validiteten af tidligere review data (30, 95, 96), hvilket gør deres sammenligninger af effektstørrelse som et potentielt upålideligt mål, navnlig fordi der ikke blev taget hensyn til indflydelsen fra finansieringskilderne i deres analyse. Dette fund stemmer overens med tidligere relateret forskning, der rapporterede ingen signifikante effekter af mobiltelefonbrug på hjernetumorer, der ofte havde industritilknytning hverken gennem finansiering eller indflydelse på studiedesign (93, 94). Samlet set tyder disse resultater på, at finansiering og forfattertilknytning har større indflydelse på studieresultaterne end studiekvaliteten.
Studier finansieret af industrigrupper, specifikke militære organisationer (især det amerikanske luftvåben) eller telekommunikationsmyndigheder var skæve i retning af at rapportere nulresultater. Det skyldes sandsynligvis de eksperimentelle metoder, der anvendes i disse studier, hvor et uforholdsmæssigt stort antal udføres som i) in vitro-eksperimenter med ii) korte eksponeringsvarigheder, iii) med simple signalmodulationer fra iv) signalgeneratorer, hvor sandsynligheden for at observere statistisk signifikante DNA-skader er meget lavere, end når der anvendes udstyr i den virkelige verden, der udsender signaler, der bærer tale eller data over længere eksponeringer. Disse designbeslutninger ser ud til at skævvride resultaterne mod nulresultater. Supplerende tabel 27 viser, hvordan studier finansieret af organisationer med særinteresser er forudindtaget mod in vitro-eksperimenter, hvilket resulterer i, at en lavere andel af studier finder effekter.
På den anden side har uafhængig finansieret forskning udført en meget bredere vifte af eksperimenter og primært rapporteret statistisk signifikante DNA-skader. Disse resultater tyder på, at de mere konservative resultater af studier af højere kvalitet sammenlignet med alle studier generelt sandsynligvis skyldes den høje andel af studier af højere kvalitet, der udføres af forskere, der har modtaget finansiering fra særinteresser, den større forekomst af in vitro-studier med korte eksponeringer fra signalgeneratorer og udelukkelsen af epidemiologiske studier på grund af dosimetribegrænsninger.
De bibliografiske netværk diagrammer i Supplerende tal 54–56 illustrerer forbindelserne mellem forskere og forskningsgrupper på dette område.
Bias om offentliggørelse
Artikler, der konkluderer, at der ikke er observeret nogen DNA-skadeeffekter, er primært offentliggjort i tre tidsskrifter: Radiation Research, Bioelectromagnetics og International Journal of Molecular Sciences. Artikler, der konkluderer, at DNA-skader er blevet fundet, vises i nogle af de samme tidsskrifter, men er mindre koncentrerede. Samlet set er uafhængig forskning blevet offentliggjort på tværs af en bred vifte af tidsskrifter. I modsætning hertil har forskning i forbindelse med industrien eller militæret gennem finansiering eller beskæftigelse i højere grad været begrænset til en lille gruppe tidsskrifter (se Tillægstabel 29); f.eks. strålingsforskning (98). Klynger af studier finansieret af særinteresser i udvalgte tidsskrifter kan forvrænge den opfattede balance mellem evidens og potentielt vildlede politiske beslutningstagere, strålesikkerhedsudøvere og offentligheden.
Generel forskningsbias
Alt det ovenstående viser, hvordan metodologiske præferencer og potentielle interessekonflikter i sagens natur påvirker resultaterne af studier (og review), hvilket reducerer robustheden af det samlede evidensgrundlag og derved skaber usikkerhed.
Dette mønster af bias er blevet dokumenteret i andre lukrative industrier, såsom lægemidler (99), landbrug og kemisk fremstilling (100). Sådanne skævheder skal anerkendes og redegøres for for at sikre integriteten og pålideligheden af offentliggjorte resultater, navnlig når der gennemføres fremtidige systematiske reviews.
Forskningens begrænsninger og huller
Begrænsninger ved dette evidenskort
Begrænsninger ved data optælling (vote counting)
Valget om at bruge optælling (vote counting) til synteser af data introducerer begrænsninger, som medfører forsigtighed ved fortolkning af resultaterne af kortlægningsprocessen.
Optælling af effektretning på tværs af primære studier er kendt for at være underberettiget, hvilket ikke giver nogen indikation af effektstørrelser eller variation af effektstørrelser i primære studier (101, 102) og ikke vægter studier efter deres præcision eller stikprøvestørrelse. Alle studier behandles ens uanset studiets størrelse, designstringens og statistiske styrke (103). Som følge heraf er de resultater, der rapporteres fra en optællingssyntese, baseret på simple optællinger af studier, der rapporterer andele af effekt vs. ingen effekt i alle dimensioner og kategorier af interesse, uanset statistisk styrke eller studiekvalitet. Resultaterne bør derfor fortolkes som informative beskrivelser af rapporterede resultater snarere end evidens for årsagssammenhæng.
Fordi optælling normalt ikke omfatter en kvalitetsvurdering af studier, er pålideligheden af den dokumentation, der udgør resultatandelene, ukendt, hvilket fører til usikkerhed med hensyn til implikationen af resultaterne. Men i den nuværende evidenskortlægningsproces blev studier kategoriseret efter tilstedeværelsen eller ej af etablerede kvalitetskriterier, som uddraget fra studieteksten. På denne måde kunne stemmeoptællinger derefter udføres for de højere kvalitetsstudier og sammenlignes med stemmeoptællinger for de mindre kvalitetsstudier. Denne proces gjorde det muligt at give større tillid til de endelige andele, der blev tildelt hver kategori, der blev undersøgt inden for opdelingen af “højere kvalitet”.
Overordnet set bør resultaterne af optællingsanalysen, der anvendes i den nuværende evidenssyntese, ses som en sonderende kortlægning, der identificerer brede mønstre langs en række dimensioner. Resultaterne kan ikke give årsags-effekt forklaringer vedrørende nogen bestemt dimension eller kategori, men de giver snarere en indikation af, hvilke dimensioner og kategorier der er mest indflydelsesrige til at vise effekter af RF-EMF på DNA-skader. Desuden indikerer resultaterne, hvor forskningshullerne er placeret for at informere fremtidig forskning.
Snævre delmængder mere velegnede til metaanalyse
Det kan hævdes, at mere præcis kvantitativ syntese kan være mulig for snævert definerede delmængder af den gennemgåede litteratur (for eksempel: en metaanalyse af studier, der bruger den samme cellelinje, eksponeringsfrekvens og kometanalyse). Sådanne homogene delmængder vil imidlertid være meget små og kan kun tilvejebringe dokumentation for meget snævre spørgsmål, hvilket ville være i modstrid med formålet med undersøgende review (scoping reviews) , dvs. at kortlægge bredden af evidens og identificere huller.
Flere forskellige dosis-respons-forhold
Et U-formet tidsdosis-respons-forhold blev postuleret ud fra pooling af alle andre dimensioner på tværs af de udpegede tidsunderkategorier (< 1 min->1 år). Nogle studier, der undersøgte eksponeringer på tværs af flere tidskategorier, gengav dog ikke denne respons; dvs. nogle viste et lineært dosis-respons-forhold (104, 105), mens andre udviste et omvendt forhold (106) eller ingen effekter (107, 108). Denne inkonsekvens kan stamme fra det begrænsede antal srudier, der anvender flere assays, der dækker mere end fire tidsbånd, så det bliver problematisk at se på individuelle studier for at validere det observerede U-formede mønster. For at bekræfte det U-formede varighedsdosis-respons-forhold og bedre forstå dets implikationer, bør fremtidig forskning inkorporere flere assays på tværs af en bredere vifte af eksponeringsintensiteter og tidsintervaller ved hjælp af standardiserede protokoller for at forbedre sammenligneligheden og reproducerbarheden af resultater.
Begrænsninger ved tidligere forskning
Mange problemer blev identificeret i både eksperimentelle og epidemiologiske studier, adskilt fra dem, der kræves til “kvalitetsvurdering”. Nogle af de vigtigere spørgsmål er opsummeret nedenfor.
Ægtheden af de signalfrie sham- eller kontrolmiljøer kunne ikke bekræftes for de fleste studier, fordi baggrundsmålinger enten ikke blev foretaget eller ikke rapporteret. Verifikation af testmiljøets EMF-integritet og identifikation af potentielle omstrejfende felter, der fungerer som forstyrrende faktorer, var derfor ikke mulig.
Mange studier viste sig at mangle vigtige metodologiske detaljer. Manglen på gennemsigtighed i metoderne gør det vanskeligt at evaluere studiekvaliteten eller at gentage studier. Sådanne studier blev klassificeret som studier af lavere kvalitet, men blev stadig inkluderet i evalueringen af DNA-skader.
Nogle studier præsenterede kun samlede data, som kan skjule potentielt “følsomme respondenter”; f.eks. viste blodet fra nogle individer mere skade ved eksponering fra andre (40–42), og lignende respons er set i millimeterbølgelængdeforskning med dyr (109, 110).
Flere epidemiologiske studier, der undersøgte mundslimhindemikrokerner fra langvarig brug af mobiltelefoner, fandt ingen statistisk signifikant forskel mellem venstre og højre kind. Forfatterne overvejede ikke ikke-lineære responser eller penetrationsevnen af RF-EMF fra mobiltelefoner. En mere relevant sammenligning er mellem tung og let brug af mobiltelefoner.
Mere specifikke problemer, der er identificeret i studier, er noteret i databasen over systematisk evidens (se Supplerende datablad 4, “Endelig studieliste (eksperimentel)”: kolonne CH og “Endelig studieliste (epidemiologisk)”: kolonne CF).
Heterogenitet af studier
En analyse af studieparametre afslører betydelig variation på tværs af studier, som blandt andet adskiller sig i eksponeringstid, intensitet, frekvenser, modulationer, analysemetoder, DNA-vurderingstidspunkter, udstyr, organisme- og celletyper. Assays bruger også forskellige pletter, forberedelsesteknikker og timinger, som kan påvirke følsomheden. Selv studier, der hævder at replikere andre, har ofte subtile forskelle, såsom variationer i dyrearter og kometanalyseparametre som beskrevet nedenfor.
Kometanalyser
En analyse af studier, der bruger kometanalysen til at måle DNA-skader, eksemplificerer yderligere heterogeniteten på tværs af studier i analyseparametre og registrering af resultater. Variationer i disse protokoller, såsom bufferformuleringer, temperatur, elektroforesespændinger, driftstider osv., kan påvirke analysens følsomhed ved at påvirke proteinfjernelse, DNA-migration og evnen til at opdage skader (34). Derudover bruger rapporterede resultater forskellige målinger (f.eks. halemoment, haleintensitet, %hale-DNA, skadeindeks osv.), som komplicerer datasyntese.
Samlet set gør denne gennemgribende heterogenitet sammen med få studier i hver kategori en kvantitativ metaanalyse til en systematisk gennemgang umulig på nuværende tidspunkt. Fremtidig forskning kræver brug af aftalte standardiserede og etablerede eksperimentelle protokoller for at forbedre repeterbarheden og sammenligneligheden på tværs af studier og for at muliggøre effektiv evidenssyntese. Dette afgrænsnings review, som behandlede over 530 studier, fremhæver det kritiske behov for en sådan standardisering.
Huller i forskningen
Evidenskortet og syntesen, der præsenteres her, har identificeret flere vigtige forskningshuller. For det første var der begrænsede studier, der undersøgte de genotoksiske effekter af RF på ikke-pattedyrarter, herunder insekter, fugle og træer. Der er også et fuldstændigt fravær af studier af krybdyr, og inden for pattedyr er der kun lidt forskning ud over studier af mennesker og gnavere.
De fleste menneskelige eksperimentelle studier anvendte forhold, der ikke afspejlede virkelige eksponeringsscenarier, typisk involverede in vitro-eksperimenter med korte, enkelte, kontinuerlige eksponeringer for et simuleret signal, der involverer en bestemt frekvens. I modsætning hertil er eksponeringer i den virkelige verden karakteriseret ved kroniske, samtidige eksponeringer for flere signaler overlejret med en række modulationsmønstre og intensiteter. Sådanne betingelser blev ikke anvendt i typiske laboratoriestudier.
Der var også få studier, der brugte højere frekvenser, såsom 5G-teknologier, på trods af deres stigende udbredelse. Forskningen halter derfor bagefter teknologiske fremskridt, hvilket efterlader potentielle risici stort set uudforskede.
Uoverensstemmelser observeret i tidligere forskning stammer fra det komplekse samspil mellem adskillige variabler. For eksempel er der identificeret både terapeutiske og potentielt skadelige effekter, hvor denne dobbelthed muligvis er knyttet til de kombinerede effekter af eksponeringens intensitet og varighed. For at afklare disse komplekse sammenhænge bør fremtidige studier metodisk adressere huller i vigtige eksperimentelle dimensioner, såsom studier af lange varigheder og ekstremt lave til meget lave eksponeringsintensiteter (se Figur 9B). Derudover har få studier undersøgt de kombinerede effekter af RF-EMF med andre miljømæssige stressfaktorer, som kan forstærke genotoksiske resultater i forskellige omgivelser og kræver yderligere studier (111).
Konsekvenser for politik og praksis
Evidenskortet viser, at mellemlang til langvarig RF-EMF-eksponering, især ved lave intensiteter, kan forårsage genetisk skade gennem ikke-termiske mekanismer såsom øget produktion af frie radikaler og oxidativ stress. Genetisk skade kan have vidtrækkende, langsigtede og potentielt irreversible konsekvenser for individuelle organismer og bredere økologisk og planetarisk sundhed (112, 113).
Både in vivo og epidemiologiske RF-EMF-studier giver troværdig evidens for genotoksicitet, hvilket tyder på potentielle risici såsom øget kræftmodtagelighed og reproduktionsskader. Studier af hjerneceller rapporterede ofte positive fund for DNA-skader, hvilket tyder på, at hjerneceller kan være særligt følsomme over for RF-EMF, hvilket indikerer en risiko for neurologiske sygdomme og hjernetumorer, som observeret i dyremodeller (114 – 116).
De nuværende retningslinjer for RF-EMF-eksponering, der er fastsat af ICNIRP (15), prioriterer forebyggelse af termiske effekter ved at inkorporere betydelige sikkerhedsmarginer (f.eks. en 50-doblet reduktion fra effekttærskler, hvor der sættes en lokal SAR-grænse på 2 W/kg for hoved og torso for den brede offentlighed, i gennemsnit over 10 gram væv). Imidlertid fandt evidenskortlægningsprocessen statistisk signifikant DNA-skade ved ekstremt lave intensiteter, hvor de laveste registrerede effekter forekom ved en SAR på 0,000000319 W/kg i et epidemiologisk studie (117) og ved 0,000003 W/kg i flere in vivo-eksperimenter (118, 119). Disse niveauer er væsentligt (>600.000 gange) under ICNIRP’s offentlige eksponeringsgrænser (15). Dette mønster antyder ikke-termiske genotoksiske effekter, fordi temperaturændringer ved disse intensiteter ville være ubetydelige og ikke målbare.
ICNIRP (2020) retningslinjer (15) fastsatte grænser for RF-EMF-eksponering for at beskytte mod termiske virkninger fra akutte eksponeringer med gennemsnitstider på 6 minutter for lokal eksponering (hoved og torso) og 30 minutter for eksponering af hele kroppen. Ovenstående analyse viste, at mellemlang (1 dag – 3 måneder) og langvarig RF-EMF-eksponering (>3 måneder eller 1.000 timer) var stærkest forbundet med genotoksiske effekter, selv ved meget lave eksponeringsintensiteter. ICNIRP’s (2020) retningslinjer (15) sætter ikke specifikke grænser for kronisk RF-EMF-eksponering på lavt niveau, især for ikke-termiske effekter som genotoksicitet, og citerer “ingen underbyggede evidens for sundhedsrelevante effekter” [(15), s. 522].
Kortlægningsprocessen afslørede også, at RF-eksponeringer er forbundet med genetisk skade i en bred vifte af organismer med en observeret følsomhed hos ikke-pattedyrsorganismer, såsom planter, insekter og muligvis padder. De nuværende retningslinjer ignorerer potentielle effekter på dyreliv eller økosystemer (78, 96). Det er værd at bemærke, at et nylig WHO-bestilt systematisk review af dyrestudier tydede på kræftfremkaldende effekter fra RF-EMF-eksponeringer (116). Andre studier tyder på biologiske effekter på ikke-menneskelige arter (120, 121). Samlet set tyder disse resultater på, at de miljømæssige konsekvenser af RF-EMF-eksponering fortjener nærmere undersøgelse (122), selvom den nuværende evidens fortsat er begrænset og omdiskuteret (96).
Selvom disse resultater endnu ikke fastslår årsagssammenhæng eller et klart niveau uden observerede negative effekter (NOAEL), indikerer de risici, som ICNIRP’s nuværende ramme udelukker ved kun at prioritere effekter med bekræftet skade [(15), s. 487]. ICNIRPs review proces og holdning beskrives bedst som en farebaseret vurdering, der kun fokuserer på bekræftede effekter. Denne tilgang er for restriktiv, da den forsinker opdateringen af retningslinjerne, indtil der er opnået absolut sikkerhed (123), hvilket muligvis ikke stemmer overens med de forsigtighedsmæssige behov for folkesundhed eller miljøbeskyttelse.
I øjeblikket er der en udbredt (6) og ofte ikke-konsensuel karakter af RF-EMF-eksponering (92) fra mobiltelefoner, basestationer og andre trådløse teknologier. Selv om man anerkender denne teknologis varighed i det moderne samfund, er der behov for politiske justeringer, der prioriterer sundhed- og miljøbeskyttelse frem for økonomiske interesser. Det kan opnås ved at anvende en forsigtighedstilgang til RF-EMF (123), og adressere potentielle risici fra ikke-termiske RF-EMF-effekter, på trods af forskningsmæssig usikkerhed. Strategier som begrundelse (vurdering af nettofordele ved RF-EMF-applikationer), optimering (balancering af beskyttelse med samfundsmæssige behov) og “Så lavt som rimeligt opnåeligt” eller “Så lavt som teknisk opnåeligt” – ALARA/ALATA (undgå deterministiske effekter og minimere stokastiske effekter) i henhold til anbefalingerne fra Den Internationale Kommission for Radiologisk Beskyttelse (ICRP) – ICRP103 (124) kan overvejes. Yderligere udvikling og implementering af trådløse teknologier bør inkorporere forbedrede sikkerhedsforanstaltninger i deres design (125), såsom at skabe enheder, der udsender lavere niveauer af RF-EMF, eller bruge materialer og antennedesign til at lede emissioner væk fra kroppen.
Derudover kan offentlig information om potentielle sundhedsrisici og personlige beskyttelsesforanstaltninger formidles gennem folkesundhedskampagner, der gør brug af eksisterende rådgivning såsom EUROPAEM EMF-retningslinjen 2016 (126); f.eks. minimering af brugen af trådløse enheder, prioritering af kablede forbindelser, opretholdelse af afstand mellem RF-EMF-kilder og kroppen, brug af luftslangeheadset eller håndfri opkald, slukning af trådløse apparater, når de ikke er i brug, og afbødning af oxidativ stress ved at inkorporere antioxidanter i kosten.
Selvom individuelle handlinger er værdifulde, er de ikke en erstatning for robuste regulatoriske standarder og ansvarlighed i branchen. Sikring af sikkerheden ved trådløse teknologier kræver en kollektiv indsats fra producenter, politikere og forbrugere for at udvikle omfattende RF-sikkerhedsretningslinjer. Fremtidige regulatoriske retningslinjer kan omfatte beskyttelsesforanstaltninger på arbejdspladsen, herunder substitution, tekniske og administrative kontroller (127), integration af bygningsbiologiske standarder (128), obligatorisk detaljeret produktmærkning for at informere brugerne om potentielle risici og standardiserede sikkerhedshygiejnepraksis.
Anbefalede handlinger
For at imødegå disse bekymringer og bygge bro over eksisterende huller anbefales følgende handlinger:
- Standardisering af forskningsprotokoller: Harmonisering af metoder på tværs af studier, især kometanalyseprotokoller, er afgørende for at reducere heterogenitet og muliggøre robuste metaanalyser.
- Fokus på langvarig og lavintensitetseksponering: Fremtidig forskning bør prioritere studier af de kumulative effekter af langvarig og lavintensitets RF-EMF-eksponering, som er mest relevante for virkelige scenarier og enheder.
- Inkludering af nye frekvenser: I betragtning af den hurtige udrulning af 5G og andre nye teknologier er der et presserende behov for forskning med fokus på højere frekvenser og nye modulationsordninger.
- Målrettede miljø- og sundhedsstudier: For at afbøde bias forbundet med industrifinansiering er større støtte til uafhængig forskning afgørende. Transparent offentliggørelse af ALLE finansieringskilder og forskertilknytninger bør være obligatorisk.
- Revurdering af RF-standarder: Reguleringsorganer skal opdatere eksponeringsretningslinjerne for at afspejle ikke-termiske mekanismer og de potentielle sundhedseffekter fra langvarige kroniske eksponeringsmiljøer ved at inkorporere resultater fra uafhængige studier af høj kvalitet.
Konklusioner
Det her præsenterede evidenskort afslører statistisk signifikant DNA-skade hos mennesker og dyr som følge af menneskeskabt RF-EMF-eksponering, især DNA-baseskade og DNA-strengbrud. Evidensen tyder også på plausible mekanistiske veje for DNA-skade, især gennem øget produktion af frie radikaler og oxidativ stress. Følsomheden over for skader varierede efter celletype, hvor reproduktionsceller (testikel, sædceller og æggestokke) sammen med hjerneceller virkede særligt sårbare. Et komplekst U-formet dosis-respons-forhold blev observeret for både eksponeringsvarighed og -intensitet, med mere DNA-skade i specifikke frekvens- og intensitetskombinationsvinduer. DNA-skader var mere tilbøjelige til at blive fundet ved hjælp af in vivo-studier, meget svage eller meget stærke signalintensiteter, meget korte eller meget lange eksponeringsvarigheder, 900, 1.800 og 2.450 MHz-frekvenser, GSM-taletilstand og pulserede moduleringer, især ved brug af virkelige enheder. På den anden side har forskning finansieret af særinteresser haft en tendens til at bruge forskellige eksperimentelle designparametre, med en høj andel af studier, der bruger in vitro, kortvarig eksponering, signaler med mellemhøj intensitet og brug af signalgeneratorer. Finansieringskilde er også en stærkere determinant for eksperimentelle resultater end kvaliteten af studiet.
Samlet set er der et stærkt evidensgrundlag, der viser DNA-skader og potentielle biologiske mekanismer, der opererer ved intensitetsniveauer, der er meget lavere end de ICNIRP-anbefalede eksponeringsgrænser. Offentlig politik kan drage fordel af implementeringen af forsigtighedsforanstaltninger såsom ALARA eller ALATA, sammen med offentlige informationskampagner for bedre at beskytte menneskers og miljøets sundhed og velvære.
Erklæring om datatilgængelighed
De oprindelige bidrag, der præsenteres i studiet, er inkluderet i artiklen/Supplerende materiale, kan yderligere forespørgsler rettes til den korresponderende forfatter.
Finansiering
Forfatteren/forfatterne erklærer, at der er modtaget økonomisk støtte til forskning og/eller udgivelse af denne artikel. SW og JM modtog økonomisk støtte til at producere denne artikel fra ORSAA (Oceania Radiofrequency Scientific Advisory Association Inc.). ORSAA er en non-profit videnskabelig forskningsorganisation, der modtager finansiering fra medlemsabonnementer og donationer uden finansiering fra industrien eller offentlige institutioner.
Anerkendelser
Forfatterne ønsker at anerkende professor Igor Belyaev for hans råd og hjælp til at forstå de teoretiske og eksperimentelle baggrunde, der er nødvendige for dette arbejde. Vi vil gerne takke Environmental Health Trust for deres generøsitet ved at sponsorere Yale 2024 Symposium, hvor indholdet af denne artikel oprindeligt blev præsenteret og er blevet tilpasset til offentliggørelse. Vi vil også gerne takke den afdøde Mr. Bruce Rowe (musiker) for hans generøse donation til ORSAA, uden hvilken denne forskning ikke ville være mulig.
Interessekonflikt
Forfatterne erklærer, at forskningen blev udført i fravær af kommercielle eller finansielle forbindelser, der kunne fortolkes som en potentiel interessekonflikt.
Generativ AI-erklæring
Forfatteren/forfatterne erklærer, at der ikke blev brugt nogen Gen AI i skabelsen af dette manuskript.
Supplerende materiale
Det supplerende materiale til denne artikel kan findes online på: https://www.frontiersin.org/articles/10.3389/fpubh.2025.1613353/full#supplementary-material
Fodnoter
1. Alle henvisninger til den supplerende tabel eller den supplerende figur henviser til detaljerne i det supplerende resultatkort (^Supplerende datablad 5).
Referencer
1. Stewart BW, Kleihues P. World Cancer Report 2003. Lyon: IARCPress (2003).
Google Scholar
2. Boyle P, Levin B. World Cancer Report. Lyon: International Agency for Research on Cancer (2008).
Google Scholar
3. Stewart B, Wild C. Wolrd Cancer Report. Geneva: WHO Press (2014).
Google Scholar
4. Wild CP, Weiderpass E, Stewart BW (editors). World Cancer Report: Cancer Research for Cancer Prevention. Lyon: International Agency for Research on Cancer (2020).
Google Scholar
5. Gu YF, Lin FP, Epstein RJ. How aging of the global population is changing oncology. Ecancermedicalscience. (2021) 15:ed119. doi: 10.3332/ecancer.2021.ed119
PubMed Abstract | Crossref Full Text | Google Scholar
6. Bandara P, Carpenter DO. Planetary electromagnetic pollution: it is time to assess its impact. Lancet Planet Health. (2018) 2:e512–14. doi: 10.1016/S2542-5196(18)30221-3
PubMed Abstract | Crossref Full Text | Google Scholar
7. IARC. Non-ionizing Radiation, Part 2: Radiofrequency Electromagnetic Fields. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, Vol. 102. Lyon: International Agency for Research on Cancer (2013).
Google Scholar
8. IARC. IARC Classifies Radiofrequency Electromagnetic Fields a Possibly Carcinogenic to Humans. Lyon (2011).
Google Scholar
9. Karipidis K, Baaken D, Loney T, Blettner M, Brzozek C, Elwood M, et al. The effect of exposure to radiofrequency fields on cancer risk in the general and working population: a systematic review of human observational studies – Part I: most researched outcomes. Environ Int. (2024) 191:108983. doi: 10.1016/j.envint.2024.108983
PubMed Abstract | Crossref Full Text | Google Scholar
10. Cheong A, Nagel ZD. Human variation in DNA repair, immune function, and cancer risk. Front Immunol. (2022) 13:899574. doi: 10.3389/fimmu.2022.899574
PubMed Abstract | Crossref Full Text | Google Scholar
11. Phillips DH, Arlt VM. Genotoxicity: damage to DNA and its consequences. EXS. (2009) 99:87–110. doi: 10.1007/978-3-7643-8336-7_4
PubMed Abstract | Crossref Full Text | Google Scholar
12. IARC. Report of the Advisory Group to Recommend Priorities for the IARC Monographs during 2025–2029. Lyon: International Agency for Research on Cancer (2024).
Google Scholar
13. FCC. RF Safety FAQ. Available online at: https://www.fcc.gov/engineering-technology/electromagnetic-compatibility-division/radio-frequency-safety/faq/rf-safety (accessed April 9, 2025).
Google Scholar
14. Dodelson S. Modern Cosmology. San Diego, CA: Academic Press (2003).
Google Scholar
15. ICNIRP. Guidelines for limiting exposure to electromagnetic fields (100 kHz to 300 GHz). Health Phys. (2020) 118:483–524. doi: 10.1097/HP.0000000000001210
PubMed Abstract | Crossref Full Text | Google Scholar
16. Panagopoulos DJ, Karabarbounis A, Yakymenko I, Chrousos GP. Humanmade electromagnetic fields: Ion forcedoscillation and voltagegated ion channel dysfunction, oxidative stress and DNA damage (Review). Int J Oncol. (2021) 59:5272. doi: 10.3892/ijo.2021.5272
PubMed Abstract | Crossref Full Text | Google Scholar
17. Vizi GN, Vandenbosch GAE. Building materials and electromagnetic radiation: The role of material and shape. J Build Eng. (2016) 5:96–103. doi: 10.1016/j.jobe.2015.11.010
Crossref Full Text | Google Scholar
18. Tayari K, Werfelli H, Chaoui M, Ghariani H, Lahiani M. Study of the effects of human tissue on performance of a loop antenna. IOSR J Elect Electron Eng. (2016) 11:06–12. doi: 10.9790/1676-1104010612
PubMed Abstract | Crossref Full Text | Google Scholar
19. Andres CMC, Lastra JMP, Juan CA, Plou FJ, Perez-Lebena E. Chemical insights into oxidative and nitrative modifications of DNA. Int J Mol Sci. (2023) 24:15240. doi: 10.3390/ijms242015240
PubMed Abstract | Crossref Full Text | Google Scholar
20. Caldecott KW. Causes and consequences of DNA single-strand breaks. Trends Biochem Sci. (2024) 49:68–78. doi: 10.1016/j.tibs.2023.11.001
PubMed Abstract | Crossref Full Text | Google Scholar
21. Abugable AA, Antar S, El-Khamisy SF. Chromosomal single-strand break repair and neurological disease: implications on transcription and emerging genomic tools. DNA Repair. (2024) 135:103629. doi: 10.1016/j.dnarep.2024.103629
PubMed Abstract | Crossref Full Text | Google Scholar
22. Shrivastav M, De Haro LP, Nickoloff JA. Regulation of DNA double-strand break repair pathway choice. Cell Res. (2008) 18:134–47. doi: 10.1038/cr.2007.111
PubMed Abstract | Crossref Full Text | Google Scholar
23. Ribezzo F, Shiloh Y, Schumacher B. Systemic DNA damage responses in aging and diseases. Semin Cancer Biol. (2016) 37–38:26–35. doi: 10.1016/j.semcancer.2015.12.005
PubMed Abstract | Crossref Full Text | Google Scholar
24. Madabhushi R, Pan L, Tsai LH, DNA. damage and its links to neurodegeneration. Neuron. (2014) 83:266–82. doi: 10.1016/j.neuron.2014.06.034
PubMed Abstract | Crossref Full Text | Google Scholar
25. Armstrong B, Baan R, Belyaev I, Benbrahim-Tallaa L, Blackman C, Blettner M, et al. Non-Ionizing Radiation, Part 2: Radiofrequency Electromagnetic Fields, Vol. 102. Lyon: International Agency for Research on Cancer (2013).
PubMed Abstract | Google Scholar
26. Belyaev I. Dependence of non-thermal biological effects of microwaves on physical and biological variables: Implications for reproducibility and safety standards. In: Giuliani L, Soffritti M, , editors. Non-Thermal Effects and Mechanisms of Interaction Between Electromagnetic Fields and Living Matter. Bologna, Italy: National Institute for the Study and Control of Cancer and Environmental Diseases “Bernardino Ramazzini”. (2010). p. 187–218.
Google Scholar
27. Lai H. Genetic effects of non-ionizing electromagnetic fields. Electromagn Biol Med. (2021) 40:1–10. doi: 10.1080/15368378.2021.1881866
PubMed Abstract | Crossref Full Text | Google Scholar
28. Ruediger HW. Genotoxic effects of radiofrequency electromagnetic fields. Pathophysiology. (2009) 16:89–102. doi: 10.1016/j.pathophys.2008.11.004
PubMed Abstract | Crossref Full Text | Google Scholar
29. Vijayalaxmi, Prihoda TJ. Genetic damage in human cells exposed to non-ionizing radiofrequency fields: a meta-analysis of the data from 88 publications (1990-2011). Mutat Res. (2012) 749:1–16. doi: 10.1016/j.mrgentox.2012.09.007
PubMed Abstract | Crossref Full Text | Google Scholar
30. Vijayalaxmi, Prihoda TJ. Comprehensive review of quality of publications and meta-analysis of genetic damage in mammalian cells exposed to non-ionizing radiofrequency fields. Radiat Res. (2019) 191:20–30. doi: 10.1667/RR15117.1
PubMed Abstract | Crossref Full Text | Google Scholar
31. Romeo S, Sannino A, Rosaria Scarfi M, Lagorio S, Zeni O. Genotoxicity of radiofrequency electromagnetic fields on mammalian cells in vitro: a systematic review with narrative synthesis. Environ Int. (2024) 193:109104. doi: 10.1016/j.envint.2024.109104
PubMed Abstract | Crossref Full Text | Google Scholar
32. Tricco AC, Lillie E, Zarin W, O’Brien KK, Colquhoun H, Levac D, et al. PRISMA extension for scoping reviews (PRISMA-ScR): checklist and explanation. Ann Intern Med. (2018) 169:467–73. doi: 10.7326/M18-0850
PubMed Abstract | Crossref Full Text | Google Scholar
33. Peters MDJ, Godfrey C, McInerney P, Munn Z, Tricco AC, Khalil H. “Scoping reviews. In:Aromataris E, , editor. JBI Manual for Evidence Synthesis: JBI. Adelaide: Joanna Briggs Institute (JBI) (2024). doi: 10.46658/JBIMES-24-09
Crossref Full Text | Google Scholar
34. Moller P, Azqueta A, Boutet-Robinet E, Koppen G, Bonassi S, Milic M, et al. Minimum Information for Reporting on the Comet Assay (MIRCA): recommendations for describing comet assay procedures and results. Nat Protoc. (2020) 15:3817–26. doi: 10.1038/s41596-020-0398-1
PubMed Abstract | Crossref Full Text | Google Scholar
35. Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. (2021) 372:n71. doi: 10.1136/bmj.n71
PubMed Abstract | Crossref Full Text | Google Scholar
36. Carlberg M, Hedendahl L, Ahonen M, Koppel T, Hardell L. Increasing incidence of thyroid cancer in the Nordic countries with main focus on Swedish data. BMC Cancer. (2016) 16:426. doi: 10.1186/s12885-016-2429-4
PubMed Abstract | Crossref Full Text | Google Scholar
37. Markovà E, Hillert L, Malmgren L, Persson BR, Belyaev IY. Microwaves from GSM mobile telephones affect 53BP1 and gamma-H2AX foci in human lymphocytes from hypersensitive and healthy persons. Environ Health Perspect. (2005) 113:1172–7. doi: 10.1289/ehp.7561
PubMed Abstract | Crossref Full Text | Google Scholar
38. Sannino A, Sarti M, Reddy SB, Prihoda TJ, Vijayalaxmi, Scarfi MR. Induction of adaptive response in human blood lymphocytes exposed to radiofrequency radiation. Radiat Res. (2009) 171:735–42. doi: 10.1667/RR1687.1
PubMed Abstract | Crossref Full Text | Google Scholar
39. Maes A, Collier M, Verschaeve L. Cytogenetic effects of 900 MHz (GSM) microwaves on human lymphocytes. Bioelectromagnetics. (2001) 22:91–6. doi: 10.1002/1521-186X(200102)22:2<91::AID-BEM1011>3.3.CO;2-N
PubMed Abstract | Crossref Full Text | Google Scholar
40. Stronati L, Testa A, Moquet J, Edwards A, Cordelli E, Villani P, et al. 935 MHz cellular phone radiation. An in vitro study of genotoxicity in human lymphocytes. Int J Radiat Biol. (2006) 82:339–46. doi: 10.1080/09553000600739173
PubMed Abstract | Crossref Full Text | Google Scholar
41. McNamee JP, Bellier PV, Gajda GB, Lavallée BF, Marro L, Lemay E. et al. No evidence for genotoxic effects from 24 h exposure of human leukocytes to 19 GHz radiofrequency fields. Radiat Res. (2003) 159:693–97. doi: 10.1667/0033-7587(2003)159[0693:NEFGEF)2.0.CO;2
PubMed Abstract | Crossref Full Text | Google Scholar
42. Waldmann P, Bohnenberger S, Greinert R, Hermann-Then B, Heselich A, Klug SJ, et al. Influence of GSM signals on human peripheral lymphocytes: study of genotoxicity. Radiat Res. (2013) 179:243–53. doi: 10.1667/RR2914.1
PubMed Abstract | Crossref Full Text | Google Scholar
43. Lai H, Levitt BB. The roles of intensity, exposure duration, and modulation on the biological effects of radiofrequency radiation and exposure guidelines. Electromagn Biol Med. (2022) 41:230–55. doi: 10.1080/15368378.2022.2065683
PubMed Abstract | Crossref Full Text | Google Scholar
44. Ghallab A. In vitro test systems and their limitations. EXCLI J. (2013) 12:1024–6. doi: 10.17877/DE290R-7558
Crossref Full Text | Google Scholar
45. Procaccini C, Pucino V, De Rosa V, Marone G, Matarese G. Neuro-endocrine networks controlling immune system in health and disease. Front Immunol. (2014) 5:143. doi: 10.3389/fimmu.2014.00143
PubMed Abstract | Crossref Full Text | Google Scholar
46. Spector NH. Interactions among the nervous, endocrine and immune systems (Nim). In: Frederickson RCA, Hendrie HC, Hintgen JN, Aprison MH, , editors. Neuroregulation of Autonomic, Endocrine and Immune Systems. Boston, MA: Springer US (1986). p. 329–41.
Google Scholar
47. Adair ER. Thermal physiology of radiofrequency radiation (RFR) interactions in animals and humans. In:Klauenberg BJ, Grandolfo M, Erwin DN, , editors. Radiofrequency Radiation Standards: Biological Effects, Dosimetry, Epidemiology, and Public Health Policy. Boston, MA: Springer US (1995):245–69.
Google Scholar
48. Robles GEH, Nelson DA. Relationship between skin temperature and blood flow during exposure to radio frequency energy: implications for device development. BMC Biomed Eng. (2025) 7:1. doi: 10.1186/s42490-024-00087-9
PubMed Abstract | Crossref Full Text | Google Scholar
49. Gapeyev A, Lukyanova N, Gudkov S. Hydrogen peroxide induced by modulated electromagnetic radiation protects the cells from DNA damage. Open Life Sci. (2014) 9:915–21. doi: 10.2478/s11535-014-0326-x
Crossref Full Text | Google Scholar
50. Sannino A, Zeni O, Romeo S, Massa R, Gialanella G, Grossi G, et al. Adaptive response in human blood lymphocytes exposed to non-ionizing radiofrequency fields: resistance to ionizing radiation-induced damage. J Radiat Res. (2014) 55:210–7. doi: 10.1093/jrr/rrt106
PubMed Abstract | Crossref Full Text | Google Scholar
51. Lai H. Levitt BB. Cellular and molecular effects of non-ionizing electromagnetic fields. Rev Environ Health. (2024) 39:519–29. doi: 10.1515/reveh-2023-0023
PubMed Abstract | Crossref Full Text | Google Scholar
52. Calvente I, Nunez MI. Is the sustainability of exposure to non-ionizing electromagnetic radiation possible? Med Clin. (2024) 162:387–93. doi: 10.1016/j.medcli.2023.11.011
PubMed Abstract | Crossref Full Text | Google Scholar
53. Jiang B, Nie J, Zhou Z, Zhang J, Tong J, Cao Y. Adaptive response in mice exposed to 900 MHz radiofrequency fields: primary DNA damage. PLoS ONE. (2012) 7:e32040. doi: 10.1371/journal.pone.0032040
PubMed Abstract | Crossref Full Text | Google Scholar
54. Sun C, Wei X, Fei Y, Su L, Zhao X, Chen G, et al. Mobile phone signal exposure triggers a hormesis-like effect in Atm(+/+) and Atm(–/–) mouse embryonic fibroblasts. Sci Rep. (2016) 6:37423. doi: 10.1038/srep37423
PubMed Abstract | Crossref Full Text | Google Scholar
55. Valavanidis A, Vlachogianni T, Fiotakis C. 8-hydroxy-2′ -deoxyguanosine (8-OHdG): a critical biomarker of oxidative stress and carcinogenesis. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. (2009) 27:120–39. doi: 10.1080/10590500902885684
PubMed Abstract | Crossref Full Text | Google Scholar
56. Wang J, Li C, Han J, Xue Y, Zheng X, Wang R, et al. Reassessing the roles of oxidative DNA base lesion 8-oxoGua and repair enzyme OGG1 in tumorigenesis. J Biomed Sci. (2025) 32:1. doi: 10.1186/s12929-024-01093-8
PubMed Abstract | Crossref Full Text | Google Scholar
57. Hsieh YC, Cheong IS, Hsu LN, Tsai HT, Tzai TS, Jou YC, et al. Clinical relevance of urinary 8-hydroxydeoxyguanosine levels in patients undergoing prostate biopsy. Mol Clin Oncol. (2024) 21:86. doi: 10.3892/mco.2024.2784
PubMed Abstract | Crossref Full Text | Google Scholar
58. Yakymenko I, Tsybulin O, Sidorik E, Henshel D, Kyrylenko O, Kyrylenko S. Oxidative mechanisms of biological activity of low-intensity radiofrequency radiation. Electromagn Biol Med. (2016) 35:186–202. doi: 10.3109/15368378.2015.1043557
PubMed Abstract | Crossref Full Text | Google Scholar
59. Jomova K, Raptova R, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, et al. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol. (2023) 97:2499–574. doi: 10.1007/s00204-023-03562-9
PubMed Abstract | Crossref Full Text | Google Scholar
60. Mortazavi SM, Mortazavi SA. Oxidative mechanisms of biological activity of low-intensity radiofrequency radiation. Electromagn Biol Med. (2016) 35:303–4. doi: 10.3109/15368378.2016.1138125
PubMed Abstract | Crossref Full Text | Google Scholar
61. Houston BJ, Nixon B, King BV, De Iuliis GN, Aitken RJ. The effects of radiofrequency electromagnetic radiation on sperm function. Reproduction. (2016) 152:R263–r76. doi: 10.1530/REP-16-0126
PubMed Abstract | Crossref Full Text | Google Scholar
62. Xie W, Xu R, Fan C, Yang C, Chen H, Cao Y. 900 MHz radiofrequency field induces mitochondrial unfolded protein response in mouse bone marrow stem cells. Front Public Health. (2021) 9:724239. doi: 10.3389/fpubh.2021.724239
PubMed Abstract | Crossref Full Text | Google Scholar
63. Chavoshani A, Aminov AR, Asgari G, Seidmohammadi A, Hashemi M. Microwave/Hydrogen Peroxide Processes. Advanced Oxidation Processes for Waste Water Treatment. Academic Press (2018). p. 215–55. doi: 10.1016/B978-0-12-810499-6.00008-5
Crossref Full Text | Google Scholar
64. Alkis ME, Akdag MZ, Dasdag S, Yegin K, Akpolat V. Single-strand DNA breaks and oxidative changes in rat testes exposed to radiofrequency radiation emitted from cellular phones. Biotechnol Biotechnol Equip. (2019) 33:1733–40. doi: 10.1080/13102818.2019.1696702
Crossref Full Text | Google Scholar
65. Sagripanti JL, Swicord ML, Davis CC. Microwave effects on plasmid DNA. Radiat Res. (1987) 110:219–31. doi: 10.2307/3576900
Crossref Full Text | Google Scholar
66. Rana JN, Mumtaz S, Choi EH, Han I. ROS production in response to high-power microwave pulses induces p53 activation and DNA damage in brain cells: radiosensitivity and biological dosimetry evaluation. Front Cell Dev Biol. (2023) 11:1067861. doi: 10.3389/fcell.2023.1067861
PubMed Abstract | Crossref Full Text | Google Scholar
67. Chemeris NK, Rubanik AV, Gapeyev AB. High power microwave pulses are not genotoxic and possess anti-inflammatory effects. In: Proceedings of 2007 17th International Crimean Conference – Microwave & Telecommunication Technology. Sevastopol (2007). doi: 10.1109/CRMICO.2007.4368615
Crossref Full Text | Google Scholar
68. Ni S, Yu Y, Zhang Y, Wu W, Lai K. Yao K. Study of oxidative stress in human lens epithelial cells exposed to 18 GHz radiofrequency fields. PLoS ONE. (2013) 8:e72370. doi: 10.1371/journal.pone.0072370
PubMed Abstract | Crossref Full Text | Google Scholar
69. Redza-Dutordoir M, Averill-Bates DA. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim Biophys Acta. (2016) 1863:2977–92. doi: 10.1016/j.bbamcr.2016.09.012
PubMed Abstract | Crossref Full Text | Google Scholar
70. McCredden JE, Cook N, Weller S, Leach V. Wireless technology is an environmental stressor requiring new understanding and approaches in health care. Front Public Health. (2022) 10:986315. doi: 10.3389/fpubh.2022.986315
PubMed Abstract | Crossref Full Text | Google Scholar
71. Sottile ML, Nadin SB. Heat shock proteins and DNA repair mechanisms: an updated overview. Cell Stress Chaperones. (2018) 23:303–15. doi: 10.1007/s12192-017-0843-4
PubMed Abstract | Crossref Full Text | Google Scholar
72. Ikwegbue PC, Masamba P, Oyinloye BE, Kappo AP. Roles of heat shock proteins in apoptosis, oxidative stress, human inflammatory diseases, and cancer. Pharmaceuticals. (2017) 11:2. doi: 10.3390/ph11010002
PubMed Abstract | Crossref Full Text | Google Scholar
73. Knowlton AA. Current concepts in transcription, translation, and the regulation of gene expression: a primer for the clinician. Chest. (1995) 107:241–48. doi: 10.1378/chest.107.1.241
PubMed Abstract | Crossref Full Text | Google Scholar
74. Dahon C, Aguida B, Lebon Y, Le Guen P, Dangremont A, Meyer O, et al. A novel method for achieving precision and reproducibility in a 1.8 ghz radiofrequency exposure system that modulates intracellular ROS as a function of signal amplitude in human cell cultures. Bioengineering. (2025) 12:257. doi: 10.3390/bioengineering12030257
PubMed Abstract | Crossref Full Text | Google Scholar
75. Clarke TL, Mostoslavsky R. DNA repair as a shared hallmark in cancer and ageing. Mol Oncol. (2022) 16:3352–79. doi: 10.1002/1878-0261.13285
PubMed Abstract | Crossref Full Text | Google Scholar
76. Li R, Ma M, Li L, Zhao L, Zhang T, Gao X, et al. The protective effect of autophagy on DNA damage in mouse spermatocyte-derived cells exposed to 1800 MHz radiofrequency electromagnetic fields. Cell Physiol Biochem. (2018) 48:29–41. doi: 10.1159/000491660
PubMed Abstract | Crossref Full Text | Google Scholar
77. Sannino A, Scarfi MR, Dufossee M, Romeo S, Poeta L, Prouzet-Mauleon V, et al. Inhibition of autophagy negates radiofrequency-induced adaptive response in SH-SY5Y neuroblastoma cells. Int J Mol Sci. (2022) 23:158414. doi: 10.3390/ijms23158414
PubMed Abstract | Crossref Full Text | Google Scholar
78. Belyaev I, Blackman C, Chamberlin K, DeSalles A, Dasdag S, Fernández C, et al. Scientific evidence invalidates health assumptions underlying the FCC and ICNIRP exposure limit determinations for radiofrequency radiation: implications for 5G. Environ Health. (2022) 21:9. doi: 10.1186/s12940-022-00900-9
PubMed Abstract | Crossref Full Text | Google Scholar
79. Ushakov VL, Borisiuk MA, Alipov ED, Smirnov SV, Beliaev I. [The use of the method of anomalous time dependence of viscosity for registration of changes in the nucleoide conformation. Part 1‘ Biofizika. (2007) 52:287–94.
Google Scholar
80. Mohan C, Das C, Tyler J. Histone and chromatin dynamics facilitating DNA repair. DNA Repair. (2021) 107:103183. doi: 10.1016/j.dnarep.2021.103183
PubMed Abstract | Crossref Full Text | Google Scholar
81. Kim SH, Kim HD, Youn B, Lee CH, Kim J. Apparent low-field microwave absorption in Escherichia coli endonuclease III: a possible way for increased DNA damage by the microwave effect. J Kor Phys Soc. (2008) 52:1065–69. doi: 10.3938/jkps.52.1065
Crossref Full Text | Google Scholar
82. Gomes AM, Orr B, Novais-Cruz M, De Sousa F, Macario-Monteiro J, Lemos C, et al. Micronuclei from misaligned chromosomes that satisfy the spindle assembly checkpoint in cancer cells. Curr Biol. (2022) 32:4240–54 e5. doi: 10.1016/j.cub.2022.08.026
PubMed Abstract | Crossref Full Text | Google Scholar
83. Garaj-Vrhovac V, Fučić A, Horvat D. The correlation between the frequency of micronuclei and specific chromosome aberrations in human lymphocytes exposed to microwave radiation in vitro. Mutat Res Lett. (1992) 281:181–86. doi: 10.1016/0165-7992(92)90006-4
PubMed Abstract | Crossref Full Text | Google Scholar
84. Koyama S, Isozumi Y, Suzuki Y, Taki M, Miyakoshi J. Effects of 245-GHz electromagnetic fields with a wide range of SARs on micronucleus formation in CHO-K1 cells. Sci World J. (2004) 4 Suppl 2:29–40. doi: 10.1100/tsw.2004.176
PubMed Abstract | Crossref Full Text | Google Scholar
85. Leszczynski D, Joenvaara S, Reivinen J, Kuokka R. Non-thermal activation of the hsp27/p38MAPK stress pathway by mobile phone radiation in human endothelial cells: molecular mechanism for cancer- and blood-brain barrier-related effects. Differentiation. (2002) 70:120–9. doi: 10.1046/j.1432-0436.2002.700207.x
PubMed Abstract | Crossref Full Text | Google Scholar
86. De Zio D, Cianfanelli V, Cecconi F. New insights into the link between DNA damage and apoptosis. Antioxid Redox Signal. (2013) 19:559–71. doi: 10.1089/ars.2012.4938
PubMed Abstract | Crossref Full Text | Google Scholar
87. McIlwain DR, Berger T, Mak TW. Caspase functions in cell death and disease. Cold Spring Harb Perspect Biol. (2013) 5:a008656. doi: 10.1101/cshperspect.a008656
PubMed Abstract | Crossref Full Text | Google Scholar
88. Nagata S, Nagase H, Kawane K, Mukae N, Fukuyama H. Degradation of chromosomal DNA during apoptosis. Cell Death Differ. (2003) 10:108–16. doi: 10.1038/sj.cdd.4401161
PubMed Abstract | Crossref Full Text | Google Scholar
89. Panagopoulos DJ, Johansson O, Carlo GL. Real versus simulated mobile phone exposures in experimental studies. Biomed Res Int. (2015) 2015:607053. doi: 10.1155/2015/607053
PubMed Abstract | Crossref Full Text | Google Scholar
90. Karipidis K, Mate R, Urban D, Tinker R, Wood A. 5G mobile networks and health-a state-of-the-science review of the research into low-level RF fields above 6 GHz. J Expo Sci Environ Epidemiol. (2021) 31:585–605. doi: 10.1038/s41370-021-00297-6
PubMed Abstract | Crossref Full Text | Google Scholar
91. Huss A, Egger M, Hug K, Huwiler-Muntener K, Roosli M. Source of funding and results of studies of health effects of mobile phone use: systematic review of experimental studies. Environ Health Perspect. (2007) 115:1–4. doi: 10.1289/ehp.9149
PubMed Abstract | Crossref Full Text | Google Scholar
92. Weller S, McCredden JE. Understanding the public voices and researchers speaking into the 5G narrative. Front Public Health. (2023) 11:1339513. doi: 10.3389/fpubh.2023.1339513
PubMed Abstract | Crossref Full Text | Google Scholar
93. Myung SK, Ju W, McDonnell DD, Lee YJ, Kazinets G, Cheng CT, et al. Mobile phone use and risk of tumors: a meta-analysis. J Clin Oncol. (2009) 27:5565–72. doi: 10.1200/JCO.2008.21.6366
PubMed Abstract | Crossref Full Text | Google Scholar
94. Prasad M, Kathuria P, Nair P, Kumar A, Prasad K. Mobile phone use and risk of brain tumours: a systematic review of association between study quality, source of funding, and research outcomes. Neurol Sci. (2017) 38:797–810. doi: 10.1007/s10072-017-2850-8
PubMed Abstract | Crossref Full Text | Google Scholar
95. Wood A, Mate R, Karipidis K. Meta-analysis of in vitro and in vivo studies of the biological effects of low-level millimetre waves. J Expo Sci Environ Epidemiol. (2021) 31:606–13. doi: 10.1038/s41370-021-00307-7
PubMed Abstract | Crossref Full Text | Google Scholar
96. Karipidis K, Brzozek C, Bhatt CR, Loughran S, Wood A. What evidence exists on the impact of anthropogenic radiofrequency electromagnetic fields on animals and plants in the environment? A systematic map protocol. Environ Evid. (2021) 10:9. doi: 10.1186/s13750-021-00252-w
PubMed Abstract | Crossref Full Text | Google Scholar
97. Simko M, Repacholi MH, Foster KR, Mattsson MO, Croft RJ, Scarfi MR, et al. Exposure to radiofrequency electromagnetic fields and IARC carcinogen assessment: Risk of Bias preliminary literature assessment for 10 key characteristics of human carcinogens. Mutat Res Rev Mutat Res. (2025). doi: 10.1016/j.mrrev.2025.108545
PubMed Abstract | Crossref Full Text | Google Scholar
98. Slesin L. “Radiation Research” and The Cult of Negative Results. Available online at: https://microwavenews.com/news-center/%E2%80%9Cradiation-research%E2%80%9D-and-cult-negative-results (accessed April 9, 2025).
PubMed Abstract | Google Scholar
99. Sismondo S. Epistemic corruption, the pharmaceutical industry, and the body of medical science. Front Res Metr Anal. (2021) 6:614013. doi: 10.3389/frma.2021.614013
PubMed Abstract | Crossref Full Text | Google Scholar
100. Bero L. Industry Influence on Research: A Cycle of Bias, in The Commercial Determinants of Health. Oxford University Press (2023). doi: 10.1093/oso/9780197578742.003.0019
Crossref Full Text | Google Scholar
101. Hedges LV, Olkin I. Vote-counting methods in research synthesis. Psychol Bull. (1980) 88:359–69. doi: 10.1037/0033-2909.88.2.359
Crossref Full Text | Google Scholar
102. Thomson H, Campbell M. Reporting Guideline for Synthesis Without Meta-analysis (SWiM). Glasgow: MRC/CSO Social and Public Health Sciences Unit, University of Glasgow; Cochrane Public Health (2020).
Google Scholar
103. McKenzie JE, Brennan SE. Chapter 12: Synthesizing and presenting findings using other methods, Vol. 6.5. London: Cochrane (2024).
Google Scholar
104. Schwarz C, Kratochvil E, Pilger A, Kuster N, Adlkofer F, Rudiger HW. Radiofrequency electromagnetic fields (UMTS, 1,950 MHz) induce genotoxic effects in vitro in human fibroblasts but not in lymphocytes. Int Arch Occup Environ Health. (2008) 81:755–67. doi: 10.1007/s00420-008-0305-5
PubMed Abstract | Crossref Full Text | Google Scholar
105. Răcuciu M. Effects of radiofrequency radiation on root tip cells of Zea mays. Rouman Biotechnol Lett. (2009) 14:4365–69.
Google Scholar
106. Sun Y, Zong L, Gao Z, Zhu S, Tong J, Cao Y. Mitochondrial DNA damage and oxidative damage in HL-60 cells exposed to 900MHz radiofrequency fields. Mutat Res. (2017) 797–9:7–14. doi: 10.1016/j.mrfmmm.2017.03.001
PubMed Abstract | Crossref Full Text | Google Scholar
107. Alessio N, Santoro E, Squillaro T, Aprile D, Briccola M, Giubbini P, et al. Low-level radiofrequency exposure does not induce changes in MSC biology: an in vitro study for the prevention of NIR-related damage. Stem Cells Cloning. (2019) 12:49–59. doi: 10.2147/SCCAA.S204166
PubMed Abstract | Crossref Full Text | Google Scholar
108. Shahin-Jafari A, Bayat M, Shahhosseiny MH, Tajik P, Roudbar-Mohammadi S. Effect of long-term exposure to mobile phone radiation on alpha-Int1 gene sequence of Candida albicans. Saudi J Biol Sci. (2016) 23:426–33. doi: 10.1016/j.sjbs.2015.05.001
PubMed Abstract | Crossref Full Text | Google Scholar
109. Shckorbatov YG, Grigoryeva NN, Shakhbazov VG, Grabina VA, Bogoslavsky AM. Microwave irradiation influences on the state of human cell nuclei. Bioelectromagnetics. (1998) 19:414–19. doi: 10.1002/(SICI)1521-186X(1998)19:7 < 414::AID-BEM2>3.0.CO;2-4
Crossref Full Text | Google Scholar
110. Radzievsky AA, Rojavin MA, Cowan A, Alekseev SI, Ziskin MC. Hypoalgesic effect of millimeter waves in mice: dependence on the site of exposure. Life Sci. (2000) 66:2101–11. doi: 10.1016/S0024-3205(00)00536-1
PubMed Abstract | Crossref Full Text | Google Scholar
111. Kostoff RN, Lau CGY. Modified health effects of non-ionizing electromagnetic radiation combined with other agents reported in the biomedical literature. In:Geddes CD, , editor. Microwave Effects on DNA and Proteins. Cham: Springer International Publishing (2017). p. 97–157.
Google Scholar
112. Jackson SP, Bartek J. The DNA-damage response in human biology and disease. Nature. (2009) 461:1071–8. doi: 10.1038/nature08467
PubMed Abstract | Crossref Full Text | Google Scholar
113. Linska Z. Understanding Genotoxic Effects: Impact on Health and Environment. J Res Rep Genet. (2024) 6:200.
Google Scholar
114. Kesari KK, Kumar S, Behari J. 900-MHz microwave radiation promotes oxidation in rat brain. Electromagn Biol Med. (2011) 30:219–34. doi: 10.3109/15368378.2011.587930
PubMed Abstract | Crossref Full Text | Google Scholar
115. Megha K, Deshmukh PS, Banerjee BD, Tripathi AK, Ahmed R, Abegaonkar MP. Low intensity microwave radiation induced oxidative stress, inflammatory response and DNA damage in rat brain. Neurotoxicology. (2015) 51:158–65. doi: 10.1016/j.neuro.2015.10.009
PubMed Abstract | Crossref Full Text | Google Scholar
116. Mevissen M, Ducray A, Ward JM, Kopp-Schneider A, McNamee JP, Wood AW, et al. Effects of radiofrequency electromagnetic field exposure on cancer in laboratory animal studies, a systematic review. Environ Int. (2025) 199:109482. doi: 10.1016/j.envint.2025.109482
PubMed Abstract | Crossref Full Text | Google Scholar
117. Ding SS, Sun P, Tian H, Huo YW, Wang LR, Han Y, et al. Association between daily exposure to electromagnetic radiation from 4G smartphone and 245-GHz wi-fi and oxidative damage to semen of males attending a genetics clinic: a primary study. Int J Clin Exp Med. (2018) 11:2821–30.
Google Scholar
118. Burlaka A, Tsybulin O, Sidorik E, Lukin S, Polishuk V, Tsehmistrenko S, et al. Overproduction of free radical species in embryonal cells exposed to low intensity radiofrequency radiation. Exp Oncol. (2013) 35:219–25.
PubMed Abstract | Google Scholar
119. Tsybulin O, Sidorik E, Brieieva O, Buchynska L, Kyrylenko S, Henshel D, et al. GSM 900 MHz cellular phone radiation can either stimulate or depress early embryogenesis in Japanese quails depending on the duration of exposure. Int J Radiat Biol. (2013) 89:756–63. doi: 10.3109/09553002.2013.791408
PubMed Abstract | Crossref Full Text | Google Scholar
120. Levitt BB, Lai HC, Manville AM 2nd. Low-level EMF effects on wildlife and plants: What research tells us about an ecosystem approach. Front Public Health. (2022) 10:1000840. doi: 10.3389/fpubh.2022.1000840
PubMed Abstract | Crossref Full Text | Google Scholar
121. Balmori A. Electromagnetic radiation as an emerging driver factor for the decline of insects. Sci Total Environ. (2021) 767:144913. doi: 10.1016/j.scitotenv.2020.144913
PubMed Abstract | Crossref Full Text | Google Scholar
122. Froidevaux JSP, Recuero Virto L, Czerwiński M, Thielens A, Park KJ. Addressing wildlife exposure to radiofrequency electromagnetic fields: time for action. Environ Sci Technol Lett. (2023) 11:3–4. doi: 10.1021/acs.estlett.3c00795
Crossref Full Text | Google Scholar
123. Gee D. Late lessons from early warnings: towards realism and precaution with EMF? Pathophysiology. (2009) 16:217–31. doi: 10.1016/j.pathophys.2009.01.004
PubMed Abstract | Crossref Full Text | Google Scholar
124. International Commission on Radiological Protection. The 2007 Recommendations of the International Commission on Radiological Protection. (2007). 1032007 p.
Google Scholar
125. Heroux P, Belyaev I, Chamberlin K, Dasdag S, De Salles AAA, Rodriguez CEF, et al. Cell phone radiation exposure limits and engineering solutions. Int J Environ Res Public Health. (2023) 20:75398. doi: 10.3390/ijerph20075398
PubMed Abstract | Crossref Full Text | Google Scholar
126. Belyaev I, Dean A, Eger H, Hubmann G, Jandrisovits R, Kern M, et al. EUROPAEM EMF Guideline 2016 for the prevention, diagnosis and treatment of EMF-related health problems and illnesses. Rev Environ Health. (2016) 31:363–97. doi: 10.1515/reveh-2016-0011
PubMed Abstract | Crossref Full Text | Google Scholar
127. Kosatsky T, Shum M, Ward HD, Gallagher RP, Anselmo F, Ross R, et al. Radiofrequency Toolkit for Environmental Health Practitioners. Vancouver, BC: British Columbia Centre for Disease Control (BCCDC) and National Collaborating Centre for Environmental Health (NCCEH) (2013).
Google Scholar
128. Maes W. Building Biology Evaluation Guidelines: Standard of Building Biology Testing Methods (SBM-2008). Rosenheim: Institute for Building Biology and Sustainability (IBN) (2008).
Google Scholar