De biologiske effekter ved de ekstremt lave frekvenser ved trådløs kommunikation
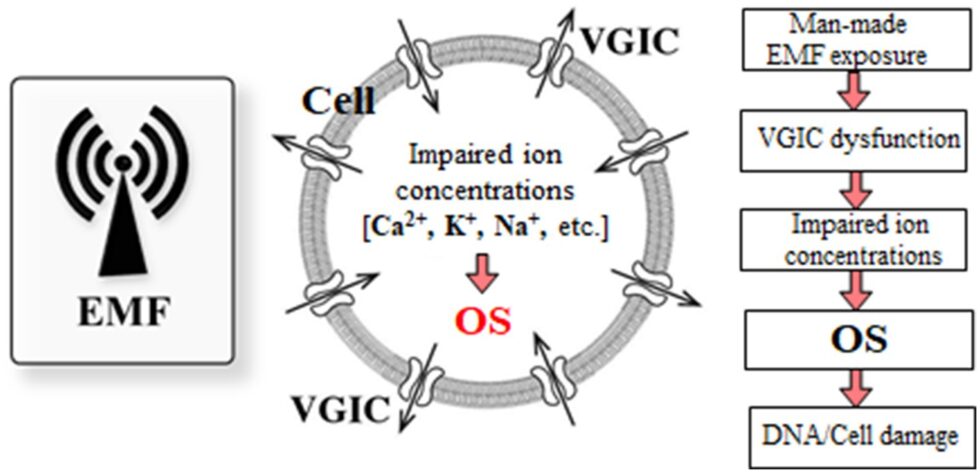
Illustration fra forskningsartiklen: Den omfattende mekanisme for menneskeskabte EMF-inducerede bioeffekter.
En vigtig forskningsartikel, der netop er udgivet af Panagopoulos et al. (2025), om hvordan elektromagnetiske felter (EMF) produceret via trådløs kommunikation er væsentligt anderledes end naturlig elektromagnetisk stråling.
Menneskeskabt trådløs kommunikation, der er polariserede, sammenhængende mikrobølger med meget variabel intensitet samt moduleret og pulseret ved ekstremt lave frekvenser, forårsager “kræft, infertilitet, elektrooverfølsomhed og forskellige andre patologier.”
Studiet er velrefereret med henvisning til 373 forskningsartikler.
“Mens menneskeskabte EMF’er ikke direkte kan ionisere molekyler, er de i stand til at gøre det indirekte i biologisk væv ved at udløse en biosyntese af reaktive iltarter (ROS), som kan beskadige biomolekyler, herunder DNA.”
Kunstigt genererede menneskeskabte trådløse kommunikationssignaler er ikke det samme som stråling fra solen, i modsætning til industriens fortælling om sikkerhed.
Studiet:
En omfattende mekanisme for biologiske og sundhedsmæssige effekter af menneskeskabte elektromagnetiske felter med ekstremt lav frekvens og trådløs kommunikation
Dimitris J. Panagopoulos1,2 Igor Yakymenko3 Geoffry N. De Iuliis4 George P. Chrousos 1,5: “A comprehensive mechanism of biological and health effects of anthropogenic extremely low frequency and wireless communication electromagnetic fields”. Frontiers (2025).
Open access: https://doi.org/10.3389/fpubh.2025.1585441
Om forskerteamet:
1) Choremeion Research Laboratory, 1st Department of Paediatrics, Medical School, National and Kapodistrian University of Athens, Athens, Greece
2) Electromagnetic Field-Biophysics Research Laboratory, Athens, Greece
3) Department of Ecology and Ecomanagement, National University of Food Technologies, Kyiv, Ukraine
4) Reproductive Science Group, School of Environmental and Life Sciences, College of Engineering, Science and Environment, University of Newcastle, Callaghan, NSW, Australia
5) University Research Institute of Maternal and Child Health and Precision Medicine and UNESCO Chair on Adolescent Health Care, National and Kapodistrian University of Athens, Medical School, Aghia Sophia Children’s Hospital, Athens, Greece
Nedenfor et større uddrag af studiet. Understregninger er tilføjet og forkortelser er nogle gange skrevet fuldt ud.
Abstrakt
Eksponeringen for menneskeskabte elektromagnetiske felter (EMF’er), især dem fra trådløs kommunikation (WC), er steget enormt. Det er et hidtil uset fænomen gennem hele den biologiske evolution, fordi alle menneskeskabte EMF’er, der er fuldt polariserede, sammenhængende og, især WC-EMF’er, meget variable, adskiller sig væsentligt fra de naturlige EMF’er. WC EMF’er består af mikrobølge (MW) bærebølger, moduleret af ELF-signaler (Extremely Low Frequency) og inkluderet i tænd/sluk-impulser, der gentages ved forskellige ELF-hastigheder. Desuden udviser de intens tilfældig variation, hovedsageligt i Ultra Low Frequency (ULF) båndet. WC EMF’er er således en kombination af mikrobølger (MW) og ELF/ULF EMF’er. Kombinationen af polarisation/kohærens og intens lavfrekvent (ELF/ULF) variabilitet ser ud til at være nøglen til EMF-bioaktivitet. Epidemiologiske studier og laboratoriestudier fremhæver en sammenhæng mellem ELF eller WC EMF eksponering og kræft, infertilitet, elektrooverfølsomhed og forskellige andre patologier. Studier finder også DNA-skader og oxidativ stress (OS), som forklarer disse patologier. Mens menneskeskabte EMF’er ikke direkte kan ionisere molekyler, er de i stand til at gøre det indirekte i biologisk væv ved at udløse biosyntesen af reaktive iltarter (ROS), som kan beskadige biomolekyler, herunder DNA. (Over)produktionen af ROS og den deraf følgende OS udløses af uregelmæssig gating af spændingsstyrede ionkanaler (VGIC’er) i cellemembranerne som beskrevet ved Ion Forced Oscillation (IFO)-VGIC-mekanismen: Mobile ioner i VGIC’er, der tvinges til at svinge af de anvendte ELF/ULF EMF’er, udøver kræfter på VGIC’ernes spændingssensorer, svarende til eller større end de kræfter, der fysiologisk blokerer disse kanaler, hvilket resulterer i deres uregelmæssige gating (dysfunktion). Dysfunktion af ionkanaler forstyrrer intracellulære ionkoncentrationer. Det udløser ROS-overproduktion og OS af de ROS-genererende systemer/enzymer i cellerne, såsom elektrontransportkæden (ETC) i mitokondrierne, eller NADPH/NADH-oxidaserne (NOX’er), nitrogenoxidsyntaserne (NOS) osv. IFO-VGIC-mekanismen og den deraf følgende OS udgør en omfattende mekanisme, der forklarer alle kendte negative biologiske og sundhedsmæssige effekter, der rapporteres at være induceret af menneskeskabte EMF’er.
1 Indledning
1.1 Unikke fysiske egenskaber ved menneskeskabte og især trådløse EMF’er: Polarisation/kohærens, kombination af frekvensbånd, modulering, pulsering og variabilitet
Alle menneskeskabte elektromagnetiske felter (EMF’er) og tilsvarende elektromagnetisk stråling (EMR) er fuldt polariserede og sammenhængende, da de produceres af elektriske/elektroniske kredsløb/antenner med specifikke geometriske former. Desuden er de fleste menneskeskabte EMF’er og især dem, der genereres af trådløse kommunikationsenheder (WC) [mobile/”smart” telefoner og tilsvarende basisantenner, trådløse husholdningstelefoner (DECT: Digitally Enhanced Cordless Telecommunications) telefoner, “wireless fidelity” (Wi-Fi) routere til trådløs internetforbindelse, “bluetooth” trådløs forbindelse mellem elektroniske enheder osv.], oscillerende og hele tiden meget variable, især i deres intensitet. Alle typer WC EMF’er består af mikrobølge (MW) bærebølger (300 MHz-300 GHz), moduleret, hovedsageligt af ekstremt lavfrekvente (ELF: 3-3,000 Hz) eller meget lavfrekvente (VLF: 3-30 kHz) signaler, og inkluderet i tænd/sluk-impulser gentaget ved forskellige ELF-hastigheder. Desuden udviser de intens tilfældig variation i deres signalamplituder med frekvenser i Ultra Low Frequency (ULF: 0-3 Hz) båndet. MW-båndet er en del af det bredere radiofrekvensbånd (RF: 300 kHz-300 GHz). Det vil sige, at selvom alle WC EMF’er normalt blot omtales som radiofrekvente (RF) EMF’er, er de faktisk en kombination af RF/MW, ELF og ULF EMF’er (1–4). Figur 1A viser 2. generations (2G) mobiltelefoni (MT) Global System for Mobile telecommunication (GSM) grundlæggende rammegentagelse, nominelt 217 Hz, pulseringer. Variabilitet i både pulsamplitude og gentagelsesfrekvens er tydelig som i alle virkelige WC-signaler. Nyere systemer 3G, 4G, 5G har grundlæggende billedgentagelsesfrekvens (nominelt) ved 100 Hz og udviser stigende variation i deres pulseringer/signaler på grund af de stadig højere mængder variabel information, de bærer (tale, billeder, video, internet osv.) (3). Figur 1B viser 100 og 200 Hz pulseringer fra en DECT-telefon.
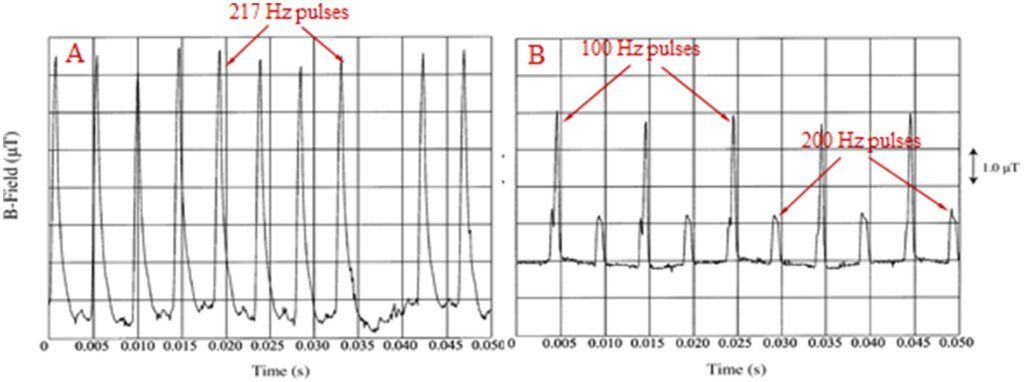
Disse unikke egenskaber gør alle menneskeskabte og mest af alle WC-EMF’er meget forskellige fra de naturlige EMF’er, som kun i specifikke tilfælde er delvist polariserede og/eller delvist i mindre grad sammenhængende (3, 5). Det geomagnetiske felt (GMF) og det geoelektriske felt (GEF) er signifikant polariserede og sammenhængende, men statiske uden signifikant variabilitet. Under magnetiske storme, som opstår cirka hvert 11. år, er der en variation på ca. 20 % i deres normale intensiteter, og derefter er der øgede forekomster af sygdom og dødelighed i menneske-/dyrepopulationerne (6).
Det ser ud til, at kombinationen af polarisation/kohærens og lavfrekvent variabilitet er nøglen til EMF-bioaktivitet. Polariserede og sammenhængende EMF’er/EMR (i modsætning til f.eks. lys og andre typer naturlige EMF’er/EMR) har ud over strålingsintensiteten netto elektriske og magnetiske felter, som udøver kræfter på alle elektrisk ladede/polære partikler/molekyler såsom de mobile ioner og de ladede/polære makromolekyler i alle biologiske systemer. Det er disse unikke egenskaber, der gør alle menneskeskabte EMF’er, og mest af alt WC EMF’er, betydeligt mere negativt bioaktive end naturlige EMF’er (3, 5, 7).
Det er gentagne gange blevet dokumenteret, at modulerede (især i amplitude) eller pulserede RF EMF’er er signifikant mere bioaktive end ikke-modulerede eller ikke-pulserende felter med samme bærefrekvens og samme intensitet som impulserne (8–29). [For reviews (se 3, 30)]. I alle tilfælde var de rapporterede effekter ikke ledsaget af nogen væsentlig opvarmning af det eksponerede biologiske væv, med andre ord var de “ikke-termiske”. Disse beviser antyder, at de ikke-termiske biologiske virkninger af WC EMF’er skyldes den inkluderede ELF-pulsation/modulering.
Derudover har ELF-EMF’er alene vist sig uafhængigt at være bioaktive, på samme måde som RF-EMF’er, der moduleres eller pulseres af ELF’er, hvilket giver yderligere bekræftelse på, at ELF-pulserings- og modulations-EMF’erne faktisk er ansvarlige for de ikke-termiske effekter af WC-EMF’er og ikke RF-bærer-EMF’erne (11, 18, 31–48). Igen var de beskrevne effekter i alle tilfælde ikke-termiske.
Dokumentationen for, at ELF/ULF og ikke RF-bærerne af de menneskeskabte/WC EMF’er er dem, der inducerer de ikke-termiske effekter, er i overensstemmelse med det faktum, at den fysiologiske elektriske aktivitet i alle former for liv er begrænset til ULF/ELF EMF’er. Der er ingen fysiologisk RF EMF i de levende organismer, heller ikke i det naturlige miljø, på trods af forvirring og misinformation blandt det videnskabelige samfund om det modsatte (6, 49–52). De såkaldte “kosmiske mikrobølger” er faktisk infrarød stråling, der når Jorden med en lavere frekvens på grund af Doppler-effekten (50, 53). Det er således tydeligt, at de ikke-termiske biologiske og sundhedsmæssige effekter, der tilskrives “RF” EMF’er, faktisk skyldes deres inkluderede ELF-pulseringer, modulation og variabilitet. Og der er praktisk talt ingen RF EMF i nogen teknisk applikation, der ikke er kombineret med ELF’er. Alle moderne digitale “RF” EMF’er indeholder ELF-pulsationer, dvs. ikke kun WC-systemer, men også radarer og radio-/tv-udsendelsessystemer (3, 4, 7, 50, 54–56). Selvom det er veldokumenteret, er det undsluppet opmærksomhed, og stadig leder forfattere efter forskellige mekanismer for ELF og “RF” EMF’er (57–59). Forfattere, der rapporterer, at de har fundet ikke-termiske EMF-effekter af ikke-modulerede og ikke-pulserede RF-bæresignaler alene, men ikke giver signalbølgeformen [som (f.eks. 60)], er enten uvidende om eksistensen af pulsationer produceret af næsten alle eksisterende RF-generatorer, eller de effekter, de rapporterer, skyldes begyndelsen og forskydningen af RF-eksponeringen (18, 56).
1.2 Menneskeskabte ELF- og WC-EMF’er: Biologiske og sundhedsmæssige effekter
Flere eksperimentelle resultater forbinder eksponering af forsøgsdyr eller celler for menneskeskabte ELF- eller WC EMF’er/EMR med oxidativt stress (OS) på grund af overproduktion af reaktive iltarter (ROS), genetiske skader/ændringer (DNA-skader, kromosomskader, mutationer osv.), cellealdring (cellealdring og tab af replikativ kapacitet), celledød og relaterede effekter [se anmeldelser i (7, 18, 43, 50, 61–66)].
Mere specifikt har talrige in vivo eller in vitro eksperimentelle laboratoriestudier vist genetiske skader og relaterede effekter induceret af menneskeskabte ELF eller WC EMF’er på en række organismer/celletyper under forskellige eksperimentelle forhold, især i de senere år. Sådanne studier er repræsentative (15, 16, 33–38, 42, 67–126). Hvis vi tilføjer studier, der fandt induktion af OS og/eller cellealdring, bliver listen meget længere (62–64, 127–133).
Flere af disse studier har fundet OS- og/eller DNA-skader med deraf følgende celledød i reproduktive celler hos forskellige dyr, hvilket resulterer i nedsat reproduktion eller embryonal død. Navnlig udviser effekterne af WC-elektromagnetiske felter på kønscellers DNA rapporteret i flere studier på en række dyr en bemærkelsesværdig lighed (68, 71, 72, 74, 75, 97–99, 107, 115, 117, 125, 130). Den genetiske skade, der findes i kønsceller, forklarer andre fund, der forbinder WC EMF-eksponering med insekt-, fugle- og pattedyrs (inklusive menneskelige) infertilitet (121, 134–144), aborter (145) eller nedgang i fugle- og insektbestande (især bier) i løbet af de seneste 20 år (146–150). Signifikant fald i reproduktion (reduceret æglægning, nedsat udvikling af reproduktive celler eller embryonal død) efter eksponering for MT-stråling blev observeret identisk hos bananfluer (97, 98, 141, 142), kyllinge- eller vagtelembryoner (71, 125, 151), oocytter fra kvæg (137), fugle (147, 148, 150) og bier (143). Lignende effekter er rapporteret for padder (152, 153), rotter og mus (107, 121, 135, 136, 138) og human sæd (nedsat antal og motilitet af sædceller) (134, 144). Disse bemærkelsesværdigt ens fund hos forskellige dyr og mennesker af forskellige forskningsgrupper kan forklares ved celledød i reproduktive celler eller embryonal død efter DNA-skader observeret i mus- eller rottesædceller (68, 107), ovarieceller fra bananfluer (72, 97–99), humane sædceller (74, 75) og vagtelembryoner (71, 125).
Det er igen bemærkelsesværdigt, at effekterne af rene ELF-EMF’er på kønsceller og reproduktion er meget lig effekterne af WC-EMF’er (31, 33, 36–38, 42, 45, 46), hvilket yderligere implicerer ELF EMF’er som et vigtigt bioaktivt stof.
Bortset fra laboratorieresultaterne om genetiske skader og infertilitet, forbinder epidemiologiske studier i stigende grad menneskeskabte ELF- eller WC EMF-eksponeringer med sundhedsproblemer, genetiske skader og kræft i menneskelige populationer. Mere specifikt er ELF EMF’er fra kraftledninger og højspændingstransformatorer (50-60 Hz) forbundet med børneleukæmi og andre kræfttyper (154–167) for magnetfeltintensiteter ned til 2 mG (0,2 μT) (159, 161) eller afstande fra elledninger på op til 600 m (157) og elektriske feltintensiteter ned til 10 V/m (155). WC EMF’er fra forskellige antenner, især radioudsendelser og MT-antenner, er blevet forbundet med forskellige former for kræft (168–171) og genetiske skader (113, 172, 173). I løbet af de sidste 15-20 år finder epidemiologiske studier en stigende sammenhæng mellem brug af mobiltelefoner eller trådløse telefoner og hjernetumorer hos mennesker (174–185). For et review af EMF-carcinogenicitetsstudier se Yakymenko og Tsibulin (171).
Desuden har andre epidemiologiske studier i løbet af de sidste 25 år fundet en sammenhæng mellem eksponering for MT/WC-antenner/-enheder med rapporterede symptomer på utilpashed kaldet “mikrobølgesyndrom” eller “elektrooverfølsomhed” (EHS). Symptomerne omfatter hovedpine, træthed, søvnforstyrrelser og forskellige andre bivirkninger (169, 186–196). En høj procentdel (~80 %) af EHS-selvrapporterende patienter blev fundet med øget OS i deres perifere blod (197). EHS-symptomer er blevet rapporteret at stige dramatisk blandt mennesker, der udsættes for 5G WC-antenner, og de omgivende RF EMF-niveauer i byer har også vist sig at stige betydeligt i løbet af de sidste 2 år efter begyndelsen af 5G-udrulningen (198, 199).
Kræft hos forsøgsdyr efter kronisk eksponering for MT/WC EMF’er er også blevet rapporteret (200, 201). Et studie af US National Toxicology Program (NTP) viste, at eksponering af rotter for simulerede 2G- eller 3G MT-emissioner (2 år, 9 timer om dagen) inducerede hjernekræft (gliom) og hjertekræft (ondartet schwannom) for både lavere og højere strålingsniveauer end de officielt accepterede grænseværdier (202). Studiet fandt også signifikant øget DNA-skade (strengbrud) i hjernen hos udsatte dyr (124), hvilket bekræfter den tætte forbindelse mellem DNA-skader og carcinogenese. En italiensk livstids eksponeringsstudie af rotter for en simuleret 2G MT EMF fandt også induktion af hjerteschwannomer og hjernegliatumorer, hvilket bekræftede resultaterne af NTP-studiet (203).
Andre studier har ikke rapporteret nogen effekter af ELF eller RF/WC EMF’er i alle ovenstående endepunkter [se reviews i (3, 18, 42, 43, 61–65, 141, 171, 204–213)], især studier, der anvendte simulerede MT/WC-eksponeringer fra generatorer med uforanderlige parametre og ingen modulering. I modsætning hertil finder mere end 95 % af de studier, der anvendte MT/WC-eksponeringer fra det virkelige liv fra kommercielt tilgængelige enheder (mobiltelefoner/trådløse telefoner, Wi-Fi osv.) med høj signalvariabilitet, effekter (3, 7, 206, 209, 210, 214–216).
Uanset virkelige eller simulerede eksponeringer finder størstedelen af de eksperimentelle studier (ca. 70 %) enten med “RF” (kombineret med ELF) eller rene ELF EMF’er effekter (62–64, 206, 208). Jagetia (64) har foretaget en omfattende gennemgang af laboratoriestudier, der omhandler genotoksiske effekter af enten ELF eller RF/WC EMF’er i en række biologiske systemer, og fandt, at blandt 207 studier fandt 144 (69,6 %) statistisk signifikante genotoksiske effekter. Langt størstedelen af de rapporterede effekter var ikke-termiske, og langt størstedelen af de anvendte EMF’er indeholdt ELF/ULF-komponenter.
Den registrerede kræftfremkaldende effekt hos mennesker og dyr, DNA/genetiske skader, OS-fund og reproduktive fald på grund af DNA-skader i æggestokke eller sædceller eller embryonal død peger alle i samme retning: Menneskeskabte EMF’er inducerer OS- og DNA-skader, infertilitet, kræft og andre relaterede patologier. Grunden til, at de samme effekter observeres hos en lang række dyr som pattedyr, fugle, insekter osv. og mennesker, er, at alle biologiske og sundhedsmæssige effekter starter i celler, og alle celler er stort set identiske i alle dyr, mennesker og endda planter. De har identiske membraner, ioner, ionkanaler og pumper, biomolekyler såsom DNA, RNA, proteiner osv., vand, ROS, identiske cellulære organeller såsom kerner, mitokondrier, ribosomer, endoplasmatisk retikulum osv., og meget lignende metaboliske processer og reguleringsmekanismer. Disse ligheder på celleniveau mellem alle dyr og mennesker er meget mere grundlæggende end forskelle i volumen, masse, form, makroskopiske funktioner, intelligens osv. Som følge heraf forventes enhver effekt induceret af EMF’er i dyreceller såsom OS, DNA-skader osv. også at blive induceret i de menneskelige celler og omvendt (7, 66).
Eksponeringsniveauerne i langt størstedelen af alle de førnævnte studier var signifikant under de officielt accepterede eksponeringsgrænser for ELF og RF EMF’er, som anbefales af den private organisation kaldet International Commission on Non-Ionizing Radiation Protection (ICNIRP) for at forhindre udladninger på mennesker i tilfælde af ELF og akut opvarmning af levende væv i tilfælde af RF/WC EMF’er. Det er bemærkelsesværdigt, at denne organisation (ICNIRP) vilkårligt ignorerer den overvældende dokumentation for ikke-termiske effekter, som udgør langt størstedelen af effekterne af menneskeskabte EMF’er, og alligevel vedtager regeringerne dens anbefalinger i stedet for at følge forsigtighedsprincippet, som dikterer det indlysende, at ingen ny teknologi bør anvendes, medmindre de, der fremmer den, har dokumenteret dens sikkerhed uden tvivl (50, 217–226).
Det Internationale Kræftforskningscenter (IARC), en afdeling i Verdenssundhedsorganisationen (WHO) har i lang tid klassificeret både ELF og “RF” (faktisk WC) EMF’er som muligvis kræftfremkaldende for mennesker (gruppe 2B) (204, 205, 227). Baseret på yderligere videnskabelig dokumentation efter IARC’s klassificering i 2011 for “RF”-EMF’er har flere studier argumenteret for, at “RF”/WC-EMF’er bør revurderes og klassificeres som sandsynligvis kræftfremkaldende (gruppe 2A) eller kræftfremkaldende (gruppe 1) for mennesker (50, 63, 64, 66, 118–120, 171, 179, 182, 183, 218, 219). Desuden har studier anmodet om en hurtig anvendelse af forsigtighedsprincippet, strengere eksponeringsgrænser, navnlig for WC EMF’s, og et moratorium for udrulningen af 5G (50, 215, 217, 218, 225, 228).
1.3 Ionisering i levende væv ved “ikke-ioniserende” EMF’er
Som det fremgår af den lange liste af laboratorie- og epidemiologiske studier, er menneskeskabte EMF-eksponeringer forbundet med OS, genetisk skade, infertilitet, EHS og kræft. Skader på DNA eller andre biologiske molekyler involverer brud på kemiske bindinger og kemiske ændringer, med andre ord ionisering. Menneskeskabte EMF’er med frekvenser op til den lave grænse for infrarød (0-3 × 1011 Hz), der undersøges her, kan ikke direkte bryde kemiske bindinger og forårsage ionisering, bortset fra meget stærke feltintensiteter (≥106 V/m) (229, 230). Sådanne feltintensiteter er sjældent til stede i miljøet, bortset fra meget tæt på højspændingsledninger og transformere eller meget tæt på atmosfæriske udladninger (lyn). Hvordan er menneskeskabte EMF’er ved miljømæssige intensiteter så i stand til at ionisere DNA og andre biologiske molekyler? Hvad er den unikke egenskab, der gør menneskeskabte EMF’er i stand til at inducere negative biologiske/sundhedsmæssige effekter i modsætning til naturlige EMF’er, herunder lys? Det har vist sig, at denne unikke egenskab er polarisering og kohærens kombineret med lavfrekvent (ULF/ELF) variabilitet (2, 3, 5, 7).
I dette arbejde giver vi en opdateret beskrivelse af, hvordan menneskeskabte EMF’er på ikke-termiske niveauer er i stand til at inducere dysfunktion af spændingsstyrede ionkanaler (VGIC’er) i cellemembraner, hvilket udløser ROS-overproduktion og OS, som igen er ansvarlig for de fleste, hvis ikke alle, kendte negative biologiske/sundhedsmæssige effekter, herunder DNA-skader og relaterede patologier. Således sker ionisering af biologiske molekyler indirekte efter menneskeskabt EMF-eksponering, medieret af den producerede ROS i cellerne (2, 210).
2 Antropogene ELF eller WC EMF’er og OS: Eksperimentel evidens
Yakymenko et al. (62) gennemgik 100 offentliggjorte eksperimentelle studier, der undersøgte OS i levende celler fra en lang række organismer (mennesker, rotter, mus, kaniner, vagtelembryoner, planter osv.), der in vitro eller in vivo blev eksponeret for RF/WC EMF’er. Af disse studier fandt 93 øget OS udtrykt som aktivering af nøgleveje, der genererer ROS-overproduktion, peroxidation, oxidativ skade på DNA, ændringer i aktiviteten af antioxidantenzymer osv. I en nyere opdatering fandt Yakymenko og Tsibulin (63), at blandt 131 offentliggjorte peer-reviewed studier, der ledte efter oxidative effekter af RF/WC EMF’er ved ikke-termiske intensiteter, i de fleste tilfælde pulseret/moduleret af ELF EMF’er, bekræftede 124 (95 %) statistisk signifikante oxidative effekter på forskellige typer biologiske systemer. Og blandt 39 offentliggjorte studier af oxidative effekter af rene ELF EMF’er fandt 36 af dem (92 %) også signifikante oxidative effekter induceret af eksponeringen. Det er derfor veldokumenteret, at menneskeskabte EMF-eksponeringer forårsager ROS-overproduktion og OS i levende celler, hvilket igen er ansvarlig for de observerede DNA-skader, infertilitet, kræft og andre relaterede patologier.
Selvom ROS på subtoksiske niveauer i cellerne fungerer som signalmolekyler involveret i forskellige fysiologiske cellulære processer, kan de også beskadige biologiske molekyler (såsom lipider, proteiner og nukleinsyrer) og forårsage forskellige sygdomme, når de er i overskud (231–234). De fleste ROS er frie radikaler. Frie radikaler er ekstremt ustabile og reaktive molekyler, der indeholder en uparret elektron betegnet med en prik (•) i deres kemiske formel. De har en meget stærk tendens til kemisk at reagere med andre molekyler og/eller med hinanden for at koble deres uparrede elektroner, balancere elektronspin og blive stabile. Denne ekstreme reaktivitet er grunden til, at de har ekstremt korte levetider. De fleste ROS reagerer hurtigt med omgivende biomolekyler og forårsager kemiske ændringer (235, 236).
To vigtige indledende frie radikaler ROS, der findes i celler efter eksponering for menneskeskabte EMF’er, er superoxidanionen (O2•−) og nitrogenoxiden (NO•) (62, 63, 125). Superoxidanionens frie radikal kan omdannes til hydroxylradikal (OH•) eller reagere med nitrogenoxid og danne peroxynitrit (ONOO−). Begge produkter (især hydroxylradikalet) er meget reaktive ROS med biologiske molekyler, især DNA (62, 125, 237, 238).
3 Biokemi i ROS
3.1 ROS-kilder i cellerne: Identitet og funktion: Afhængighed af ionkoncentrationer
3.1.1 Mitochondria
(….)
3.1.2 NADPH/NADH oxidaser (NOXs)
(….)
3.1.3 Nitrogenoxid syntaser (NOS)
(….)
3.1.4 Xanthinoxidase (XO)
(….)
3.1.5 Andre ROS-genererende/fremmende enzymer
(….)
3.2 ROS-effekt på DNA og andre biologiske molekyler
(….)
3.2.1 Peroxynitrit
(….)
3.2.2 Hydroxylradikal
(….)
4 Menneskeskabte EMF’er og VGIC’er (Spændingsstyret ionkanal)
4.1 VGIC’er: De mest følsomme elektromagnetiske sensorer i levende organismer
Tidligere studier har antaget eksistensen af specifikke elektro/magneto-sensororganer/celler i dyr/mennesker for at forklare de biologiske effekter af EMF’er [se review i (297)]. Det er ikke nødvendigt, da alle celler i alle dyr, herunder mennesker og endda planter, er udstyret med de mest følsomme EMF-sensorer, som ikke er andre end VGIC’erne (spændingsstyrede ionkanaler) den mest udbredte klasse af ionkanaler i alle cellemembraner (231, 234, 271, 297, 298).
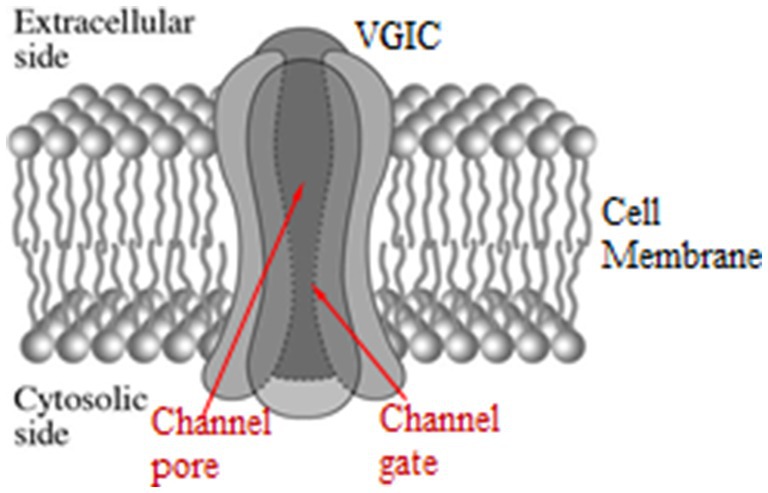
Normalt konverterer VGIC’er mellem åben og lukket tilstand ved membranspændingsændringer dV ≥ 30 mV, som udøver kræfter på deres spændingssensorer. Mere specifikt reagerer VGIC’er på ændringer mellem \u201230 og \u2012100 mV. VGIC’ernes spændingssensorer er fire symmetrisk arrangerede, transmembrane, positivt ladede parallelle α-helixer (underenheder), hver med navnet S4. De indtager den 4. position i en gruppe på 6 parallelle α-helixer (S1-S6), og er de tætteste helixer til porerne bortset fra S5-S6-helixerne, som danner porevæggene. Kanalen består af fire identiske sådanne grupper (hovedenheder I-IV) i symmetriske positioner omkring kanalens pore (Figur 2). Sensorerne er positive Lys- og Arg-aminosyrer i S4-spiralerne. Den effektive (netto)ladning på hver S4 er beregnet til at være q = 1,7qe, hvor qe er den elementære ladning. De positive ladninger fra S4-sensorerne er parret med negative ladninger fra tilstødende spiraler, så nettoladningen på porens vægge er nul. Ionerne passerer dehydreret og i en enkelt fil gennem kanalporten, den smalleste del af poren (Figur 2). Mindst fire dehydrerede mobile ioner befinder sig meget tæt på S4-sensorerne i en afstand på mindre end 1 nm, da mindst tre andre er bundet på specifikke ionbindingssteder meget tæt på porten, bortset fra den eller de ioner, der kan passere gennem porten når som helst eller er lige uden for porten klar til at passere (234, 297, 299–304).
4.2 Antropogene EMF’er og VGIC-dysfunktion: IFO-VGIC-mekanismen
En biofysisk mekanisme for EMF-inducerede biologiske effekter er blevet beskrevet i Panagopoulos et al. (2, 5, 297, 305, 306) og Panagopoulos (302). Den forklarer, i standardfysik og biologi, hvordan polariserede, sammenhængende og langsomt varierende (ULF/ELF/VLF) EMF’ER, SELV VED MEGET LAVE FELTINTENSITETER, KAN FORÅRSAGE UREGELMÆSSIG GATING (ÅBNING/LUKNING) AF VGIC’ER I CELLEMEMBRANER MED DERAF FØLGENDE FORSTYRRELSE AF CELLENS ELEKTROKEMISKE BALANCE, REDOXTILSTAND OG HOMEOSTASE. DA ELF/ULF/VLF EMF’ER ER GRUNDLÆGGENDE KOMPONENTER I TRÅDLØSE (WC) EMF’ER, TEGNER DENNE MEKANISME SIG FOR LANGT STØRSTEDELEN AF DE BIOLOGISKE EFFEKTER AF ALLE MENNESKESKABTE (POLARISEREDE, SAMMENHÆNGENDE OG VARIERENDE) EMF’ER.
Mens VGIC’er normalt er styret af \u201230-100 mV spændingsændringer i det meget stærke transmembranfelt, med andre ord reagerer på feltændringer mellem 3 × 106 og 107 V/m, kan de også reagere på meget svagt polariserede, sammenhængende og langsomt varierende EMF’er ned til \u201210-5 V/m via den tvungne svingning, som EMF’er inducerer på mobile ioner i umiddelbar nærhed (<1 nm) til sensorerne (ion-forced-oscillation: IFO). Det sker, fordi den kraft, der udøves på S4-sensorerne af oscillerende ioner i umiddelbar nærhed, afhængigt af den omvendte tredjedels potens af afstanden mellem ladninger (se Ligning 12 nedenfor), er meget større end en direkte kraft fra en eksternt påført EMF, som afhænger af den første potens af det anvendte felt (2, 5, 297). De førnævnte (mindst) fire ioner tæt på poreporten, når de er tvunget til at svinge parallelt og i fase, udøver konstruktive koordinerede kræfter på S4-sensorerne, der er i stand til at porte kanalen (Figur 3).
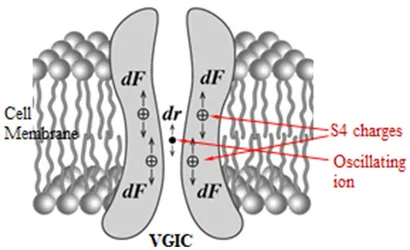
Kræfter, der udøves af en ekstern polariseret EMF på en hvilken som helst mobil ion inden for en VGIC, forårsager en forskydning dr af ionen fra dens “oprindelige” position, som igen udøver en yderligere Coulomb-kraft dF på S4-spændingssensorerne på VGIC’en, hvilket kan resultere i åbning/lukning (port) af kanalen (2, 5, 297, 305, 306) (Figur 3).
(….)
(….)
5 VGIC-dysfunktion, der fører til OS: Forbinder prikkerne for en omfattende mekanisme for EMF-inducerede biologiske og sundhedsmæssige effekter
Hvordan kan den indledende ROS (O2•− og NO•), der genereres efter EMF-eksponering, produceres af VGIC-dysfunktion? Dette var et missing link indtil for nylig, hvor vi specifikt ledte efter sådan dokumentation (2, 210). Vi indså, at selvom masser af data, der forbinder nedsat ionkanalfunktion og induktion af celledød eller kræft, havde været tilgængelige i lang tid (266, 267), og selv om de fleste ionkanaler, især kationkanaler, er VGIC’er, er forbindelsen mellem VGIC-dysfunktion induceret af EMF-eksponering og OS (2, 210, 238, 319–322), der førte til DNA/cellulær skade, undsluppet den nødvendige opmærksomhed.
Mange studier har fundet en sammenhæng mellem Ca2+, K+, Na+ og Cl− VGIC dysfunktion med OS og relaterede patologier (238, 319, 321, 322). Det er gentagne gange vist, at VGIC-dysfunktion induceret af menneskeskabte EMF’er kan udløse øjeblikkelig ROS-generering i cellerne, med denne effekt signifikant formindsket ved brug af specifikke ionkanalblokkere (238, 259, 314, 316). Nyere forskning bekræfter yderligere sammenhængen mellem VGIC-dysfunktion og ROS (over)produktion. For eksempel blev ROS-overproduktion gennem aktivering af NADPH-oxidase af ekstracellulært tau-protein i co-kulturer af neuroner og astrocytter reduceret i nærvær af nifedipin, hæmmer af Ca2+ VGIC (323). Epitelcelledød forbundet med forhøjelse af ROS blev forhindret af lidocain, en velkendt Na+ VGIC-hæmmer med antioxidantvirkninger (324). Induceret ROS-produktion i murin mikroglia blev hæmmet på en dosisafhængig måde af K+ VGIC-blokering og i mere begrænset grad af Cl−kanalblokering (325).
Forskellige patologiske tilstande, herunder neurodegenerative sygdomme, kaldet “kanalopatier”, er blevet fundet som værende forårsaget af ionkanaldysfunktion. Svækkelse af enten spændings- eller ligandstyrede ionkanaler er blevet identificeret som en årsag til neurologiske sygdomme. De involverede ionkanaler omfatter Ca2+, K+ og Na+ VGIC’er (318, 326). Flere studier har dokumenteret sammenhængen mellem Ca2+, K+, Na+ og Cl− kanaldysfunktion og udviklingen af OS-relaterede patologier (321). Ionkanaldysfunktion, der fører til OS, er også en almindelig årsag til degenerative sygdomme i centralnervesystemet (CNS) af forskellige genetiske ætiologier og er en almindelig faktor i neurologiske lidelser. Ionkanalernes rolle i neurodegenerative lidelser forbundet med OS er nu blevet anerkendt, da ionkanalerne gennemgår funktionelle justeringer under sådanne forhold (318, 327).
Det er tydeligt, at funktionen af ionpumper og kanaler styrer de intracellulære koncentrationer af mobile ioner, og igen funktionen af de cellulære systemer/enzymer, der producerer ROS (2, 210). Enhver dysfunktion i ionkanaler vil påvirke de ellers omhyggeligt kontrollerede intracellulære ionkoncentrationer, hvilket forstyrrer cellens elektrokemiske balance og homeostase, herunder den intracellulære redoxstatus, som er et indeks for ROS-indholdet i cellen. Af den dokumentation, der er fremhævet her, følger det, at forstyrrelse af ionhomeostase kan udløse OS ved ROS-overproduktion og efterfølgende DNA-skade. Omvendt kan den intracellulære redoxstatus ændre gating-egenskaberne for ionkanaler og udløse åbning eller lukning af Ca2+-, Na+– og K+-kanaler for at genindføre homeostase (318, 331). Ionkanaler er således portvogtere af redoxstatus, og cellens elektrokemiske balance og homeostase (305, 328).
For eksempel er Ca2+ en kritisk signalfaktor, der regulerer mange cellefunktioner, herunder celleproliferation, differentiering og apoptose (233, 244, 267, 329). Ændringer i intracellulære Ca2+ niveauer afkodes af Ca2+ -sensorer, som initierer signalering for forskellige fysiologiske processer (330). Ændringer i Ca2+ homøostase og signalering er ofte forbundet med forskellige patologiske tilstande, herunder kræft. ROS-reguleringssystemet er tæt forbundet med Ca2+-signalsystemet, som fungerer ved ændringer i intracellulære Ca2+-koncentrationer. Dysfunktion af Ca2+ kanaler i plasmaet eller mitokondriemembranen vil forstyrre signalsystemet og øge ROS-niveauerne i enhver celle, hvilket forstærker skadelige virkninger, herunder cytotoksicitet og resulterer i patogenese af forskellige lidelser (44, 244, 321, 322, 328, 330, 331). Omvendt kan ROS påvirke calciumkoncentrationen i cellen betydeligt ved at ændre funktionen af Ca2+ kanaler (233).
Øgede niveauer af intracellulær Ca2+ er i nogle tilfælde forbundet med øget apoptose, sandsynligvis på grund af aktivering af Ca2+ afhængig DNase I (332). Dette kan være en alternativ vej for DNA-skader og relaterede patologier. Ændringer i normale Ca2+ niveauer i mitokondrierne kan inducere frigivelse af cytokrom C, et mitokondrieprotein, som er et signalmolekyle for apoptose i cytoplasmaet, som derefter fortsætter med at initiere apoptose i cellen, og aktivering af nukleaser, som vil forårsage DNA-skade (330).
Effekten af menneskeskabte EMF’er, især ELF, eller RF pulseret eller amplitudemoduleret af ELF-signaler, på calciumkoncentrationer i eksponerede celler og den unikke rolle, som calcium-VGC’erne eller VGCC’erne spiller i EMF-inducerede bioeffekter, har været veldokumenteret i lang tid (8–10, 18, 22, 44, 238, 298, 314–316, 333–341), og forklaret ved hjælp af IFO-VGIC-mekanismen (2, 5, 297, 305, 306). Dysfunktion af VGCC’er vil forårsage ændringer i de intracellulære calciumkoncentrationer, svækkelse af Ca2+-signalsystemet og deraf følgende ROS-overproduktion.
Walleczek (44) gennemgik mange studier, der viser effekter af ELF EMF’er på celler i immunsystemet, hvilket afslører den kritiske rolle af intracellulært calcium. Men indtil da var stedet for interaktion mellem EMF’er og celler ukendt, selvom fakta pegede på calciumionkanalerne i cellemembranerne som en yderst rimelig forklaring. Samtidig har Liburdy (315) i en række banebrydende eksperimenter vist, at calciumtilstrømning i lymfocytter, som opstod inden for få minutter efter begyndelsen af ELF EMF-eksponeringer, skyldtes en effekt på calciumkanalerne i celleplasmamembranerne (hvoraf de fleste er spændingsstyrede) og ikke på grund af frigivelse fra intracellulære lagre.
Bortset fra effekten af EMF’er på Ca2+, Na+, K+ osv. VGIC’er, vil proton (H+) VGIC’er blive påvirket på samme måde (342, 343). Det vil igen forstyrre funktionen af NOX’er, der udløser ROS-generering (punkt 3.1.2). Således er ikke kun VGCC’er, men alle VGIC’er de steder, hvor virkningerne af menneskeskabte EMF’er på celler finder sted (2, 210).
Udover mange andre bivirkninger kan ROS også påvirke ionkanalerne selv. For eksempel indeholder mange ionkanaler cysteinrester med meget reaktive thiolgrupper (SH). Disse er særligt modtagelige for oxidation af ROS. Oxidation af cysteinrester kan føre til dannelse af disulfidbindinger. Det kan ændre kanalkonformationen og påvirke kanalporten. En anden effekt af ROS på VGIC’er kan være dannelsen af sulfensyre, sulfinsyre eller sulfonsyre: Disse modifikationer kan ændre kanalens struktur og funktion, hvilket potentielt kan føre til kanalinaktivering eller ændret ionpermeabilitet (318). Oxidation af K+-kanaler med ROS er en almindelig begivenhed i den aldrende hjerne (344). Derfor fører ionkanaldysfunktion til ROS-overproduktion, og ROS forstærker ionkanaldysfunktionen yderligere. Det er klart, at vi har en ond cirkel her, hvor VGIC-dysfunktion fører til OS i celler, hvilket igen forstyrrer ionkanalerne endnu mere, og fører til endnu mere udtalt OS.
Balancen mellem de forskellige mobile ioner i en celle er tæt forbundet med og bestemmer faktisk cellens homeostase. ROS-produktion i alle celler starter efter ubalance i ionkoncentrationer. Dysfunktion af ionkanaler eller pumper af en eller anden grund, herunder EMF-eksponering, kan let forårsage ionisk ubalance, ROS-overproduktion og OS. Figur 5 viser de biokemiske processer relateret til OS, og initieret efter EMF-induceret dysfunktion af VGIC’er og ubalance i ionkoncentrationer.
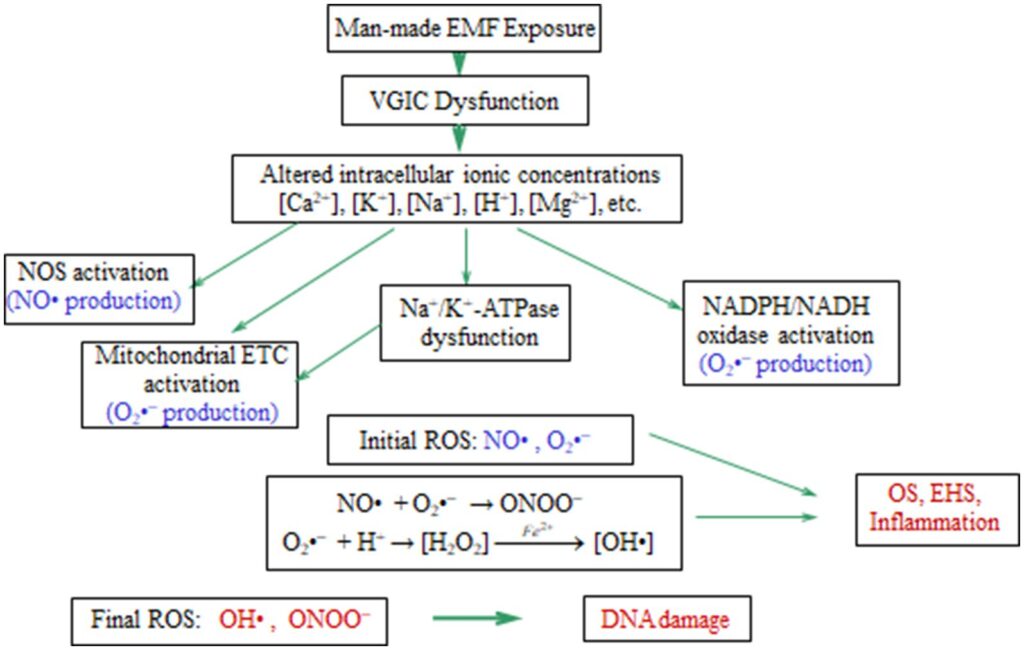
Det er således veldokumenteret, at ionkanaldysfunktion forårsager OS, og her argumenterer vi for, at OS fundet efter menneskeskabt og især WC EMF-eksponering induceres efter VGIC-dysfunktion. Vi har en klar rækkefølge af begivenheder, der starter fra uregelmæssig gating af VGIC’er af menneskeskabte EMF’er op til OS, celle/DNA-skader og relaterede patologier, herunder infertilitet og carcinogenese. Derfor kan en omfattende mekanisme for EMF-inducerede bioeffekter klart afgrænses, med et biofysisk stadium, der forårsager VGIC-dysfunktion og ionisk ubalance, og et efterfølgende biokemisk stadium, der resulterer i OS-relateret patogenese.
Figur 6 viser en skematisk repræsentation af den beskrevne omfattende mekanisme initieret af EMF-induceret dysfunktion af VGIC’er og resulterer i OS og cellulær skade.
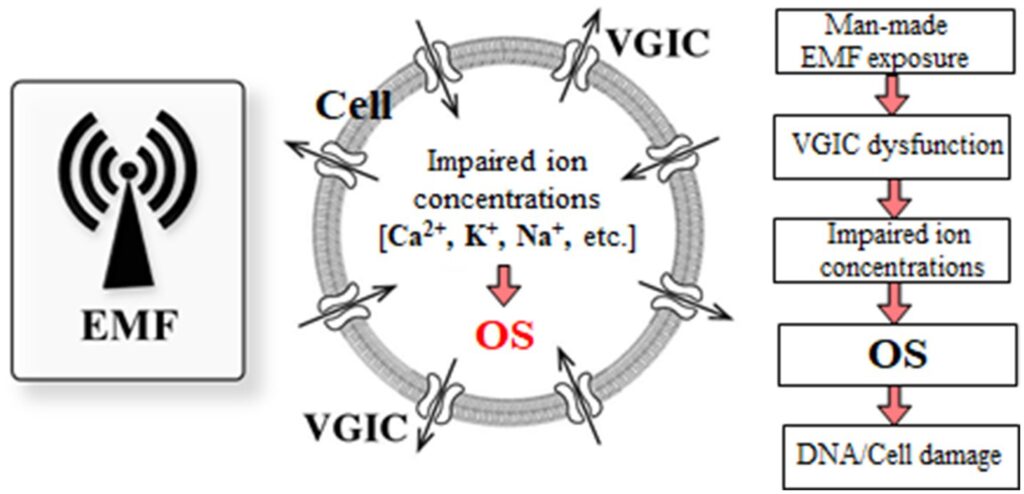
6 Diskussion
Vi har gennemgået eksperimentelle og epidemiologiske studier, der refererer til de biologiske og sundhedsmæssige effekter af menneskeskabte ELF- og WC EMF-eksponeringer. Vi finder endnu en gang, at det er veldokumenteret, at både rent ELF og WC/RF (indeholdende ELF) menneskeskabte EMF’er inducerer OS og genetisk skade, hvilket kan føre til relaterede patologier, såsom infertilitet og kræft hos både mennesker og dyr.
Vi har dokumenteret, at alle menneskeskabte EMF’er omtalt som “RF”, især WC EMF’er, bortset fra deres RF-emissioner (bærebølger), udsender ELF/ULF/VLF EMF’ER I FORM AF MODULERING, PULSERING OG VARIABILITET, OG DERMED FAKTISK ER EN KOMBINATION AF RF OG ELF/ULF/VLF EMF’er.
Nogle forfattere forveksler “pulsering” med periodisk eksponering (intermitterende). Zahumenska et al. (373) anvendte en intermitterende eksponering (6 × 10 minutter) for en kontinuerlig bølge LF EMF (87-207 kHz) med 10 minutters pauser mellem eksponeringsperioderne og hævdede, at de undersøgte effekterne af pulserende EMF og ikke fandt nogen signifikant forskel fra fraværet af effekter ved en uafbrudt eksponering (1 × 60 min.). Det er ikke tilfældet. I et “pulserende felt” er tænd/sluk-pulseringerne iboende i signalet og forekommer ved ELF/VLF-hastigheder, normalt i størrelsesordenen hundreder/tusinder pulseringer i sekundet, mens feltet/signalet i den “intermitterende” eksponering som i dette tilfælde afbrydes eksternt af en timer eller endda manuelt ved hjælp af en kontakt. Mens pulserende EMF’er i næsten alle tilfælde viser sig at producere betydeligt større effekter end kontinuerlige (ikke-pulserede) EMF’er, kan en intermitterende eksponering for enhver EMF give mindre effekter end en uafbrudt eksponering for den samme EMF, når intermittensintervallet er langt nok (f.eks. ≥10 min) til at tillade den eksponerede organisme at reparere skader og/eller tilpasse sig stressfaktoren (72). Ved at forveksle pulsering med intermittens kan man drage helt vildledende konklusioner. Zahumenska et al. (373) fandt faktisk ingen effekt ved brug af en kontinuerlig bølge LF EMF, som var forventet, men de hævdede, at de ikke fandt nogen effekt med en “pulserende” EMF. En definition af de forskellige fysiske parametre for EMF’er kan findes i Panagopoulos og Margaritis (345) og Panagopoulos et al. (3).
Vi har forklaret, at alle menneskeskabte EMF’er er fuldt polariserede og sammenhængende, med lavfrekvente (ELF/ULF/VLF) intensitetsvariationer i langt de fleste tilfælde, hvilket betyder, at de har netto elektriske og magnetiske felter, der svinger (ved ELF/ULF/VLF-hastigheder) i enkeltretninger og i fase. Denne tilstand inducerer parallelle og sammenhængende lavfrekvente tvungne svingninger af mobile ioner og andre ladede/polære molekyler i levende væv. IFO-VGIC-mekanismen har beskrevet, hvordan sådanne svingninger inducerer dysfunktion af VGIC’er i membranerne i alle celler, hvilket resulterer i ændrede intracellulære ionkoncentrationer (2, 5, 297, 305, 306).
Ifølge IFO-VGIC-mekanismen induceres de ikke-termiske biologiske/sundhedsmæssige effekter, der er rapporteret i litteraturen, specifikt af lavfrekvente (ULF/ELF/VLF) og ikke de højfrekvente (rent RF/MW), EMF’er. Det forklarer, hvorfor de ikke-termiske effekter, der før tilskrives “RF” EMF’er, forsvinder i fravær af lavfrekvent modulation/pulsering/variabilitet. Det følger heraf, at rene RF/MW EMF’er kun kan inducere opvarmning af biologisk væv ved tilstrækkeligt høje intensiteter og frekvenser, der nærmer sig infrarød (2, 3). En oversigt over VGIC’s struktur og funktion samt IFO-VGIC-mekanismen er givet i afsnit 4.
Det er vigtigt at bemærke, at VGIC’er ikke er styret af direkte kræfter på deres S4-sensorer af eksternt påførte EMF’er. Det ville kræve påførte felter i størrelsesordenen af de transmembranfelter (~107 V/m) (56). Grunden til, at selv meget svage (ned til 10−5–10−4 V/m) ULF/ELF-menneskeskabte felter kan portere VGIC’er, er, at de på grund af deres polariserede og sammenhængende karakter kombineret med lavfrekvent variabilitet kan inducere parallelle og koordinerede lavfrekvente tvungne svingninger af mobile ioner i kanalerne. Og de kræfter, der udøves på S4-sensorerne af flere oscillerende ioner i umiddelbar nærhed (≤1 nm), afhængigt af afstandens omvendte tredje potens (Ligning 12), er meget større end direkte kræfter fra eksternt anvendte elektromagnetiske felter. Med andre ord, på grund af IFO-fænomenet i umiddelbar nærhed af VGIC-sensorerne, forstærkes kræfterne enormt. Det er et nøglepunkt i forståelsen af IFO-VGIC-mekanismen.
Det er således polarisering og kohærens kombineret med lavfrekvent variabilitet, der gør menneskeskabte EMF’er i stand til uregelmæssigt at portere (åbne eller lukke) VGIC’er, de mest følsomme EMF-sensorer og den mest udbredte klasse af ionkanaler i alle cellemembraner i alle levende organismer. Det forårsager forstyrrelse af ionkoncentrationer i cellerne, hvilket igen udløser (over)produktion af ROS. ROS kan let forårsage ionisering/kemiske ændringer i levende væv, dvs. brud på kemiske bindinger og DNA-skader.
Vi beskrev biokemiske processer initieret i levende celler ved dysfunktion af VGIC’er på grund af menneskeskabt EMF-eksponering, hvilket fører til ændrede intracellulære koncentrationer af kritiske ioner såsom Ca2+, Na+, K+, H+ osv., og forstyrrelse af cellens elektrokemiske balance, redoxtilstand og homeostase. Det fører til øjeblikkelig produktion af de to indledende ROS, superoxidanion (O2•−) og nitrogenoxid (NO•), som derefter let kan omdannes til den kraftige “endelige” ROS peroxynitrit (ONOO−) og/eller hydroxylradikal (OH•), som kan beskadige DNA eller ethvert andet biologisk molekyle.
Det er bemærkelsesværdigt, at den samme “endelige” ROS, der i sidste ende forårsager biologisk skade i tilfælde af EMF’er (“ikke-ioniserende” strålinger), hydroxylradikal og peroxynitrit, også findes i tilfælde af eksponering for ioniserende stråling. Det anslås, at omkring 2/3 af DNA-skaderne forårsaget af ioniserende stråling skyldes OH• (276, 294, 295, 346–348). Det giver et svar på påstande om, at “ikke-ioniserende” menneskeskabte EMF’er umuligt kan forårsage biologisk skade. Det kommer sådan, at den samme ROS, der faktisk udfører den biologiske skade i de fleste tilfælde, produceres af enten ioniserende stråling eller “ikke-ioniserende” EMF’er/EMR. Det hænger sammen med det faktum, at strålingens virkning i biologisk væv i de fleste tilfælde er indirekte. Det eksterne middel forårsager svækkelse af cellehomeostase, og som svar genererer cellen ROS, som udfører skaden.
ROS-kilder i celler er ETC i mitokondrierne, ETC i NOX’erne i plasmamembranen, NOS-enzymerne forskellige steder i cellen og forskellige andre sekundære kilder (beskrevet i pkt. 3.1). Alle ROS-kilder/promotorer påvirkes af de intracellulære koncentrationer af kationer som Ca2+, K+, Na+, H+, hvor de fleste kationkanaler er spændingsstyrede (VGIC’er) (231, 234, 271). Derfor kan alle ROS-kilder i celler blive påvirket af menneskeskabte EMF’er. Det er bemærkelsesværdigt, at i alle tilfælde, der hidtil er rapporteret i litteraturen, øger og ikke reducerer ROS/OS i celler. Det er en yderligere indikation af, at cellerne opfatter menneskeskabte EMF’er som en forstyrrelse.
Selvom mange af detaljerne i ionsignaleringen, der udløser ROS-generering af ovenstående kilder, stadig er uudforskede, ved vi, at udløsningen involverer ændringer i de intracellulære ionkoncentrationer. Da menneskeskabte EMF’er har evnen til at forårsage dysfunktion af VGIC’er, er de grundlæggende dele af hele processen, der fører til DNA-skader og relaterede patologier, allerede identificeret, og prikkerne er allerede forbundet, hvilket afslører den komplette EMF-inducerede bioeffektmekanisme.
Der er stadig flere spørgsmål, der skal behandles. For eksempel diskuterede vi ikke tilstanden af antioxidantsystemet (AOS) under tilstanden af kronisk OS på grund af langvarig EMF-eksponering. Da produktionen af ROS på fysiologiske niveauer er en væsentlig del af enhver celles liv, er AOS’s rolle at begrænse niveauet af ROS under OS-tærsklen, hvor der vil opstå skader. Desuden kontrollerer AOS aktiviteten af reparationsenzymer. Celler/organismer med kompromitteret antioxidantkapacitet eller høje energibehov er særligt sårbare over for OS og efterfølgende over for menneskeskabte EMF’er. Mange studier har afsløret signifikante ændringer i aktiviteten af vigtige antioxidantenzymer under moduleret og/eller pulseret RF/WC EMF-eksponering [se reviews i (62–64)]. Og mens ændringerne i antioxidantenzymers aktiviteter i mange tilfælde kan induceres af ROS-overproduktion i de eksponerede celler, kan de også blive påvirket af ioniske ubalancer relateret til VGIC-dysfunktion. For eksempel kan Ca2+ påvirke aktiviteten af transkriptionsfaktorer som NF-KB og Nrf2, som regulerer ekspressionen af antioxidantgener (349). Yderligere kan forstyrrelse af Na+/K+ gradienter indirekte påvirke Ca2+ homeostase gennem Na+/ Ca2+ veksleren, som kan fungere i begge retninger afhængigt af iongradienterne (350).
Når overproduktion af ROS i en celle overstiger kapaciteten af dens AOS, er cellen/organismen under OS. En vedvarende eller gentagen sådan tilstand fører til DNA/cellulær skade. Intracellulære ioner, især Ca2+, påvirker aktiviteten af AOS og DNA-reparationsenzymer. For eksempel er nogle DNA-reparationsveje Ca2+-afhængige. Forstyrrelse af Ca2+ homeostase kan derfor forringe DNA-reparationskapaciteten, hvilket gør celler mere modtagelige for DNA-skader (351). Ureparerede/fejlreparerede DNA-læsioner såsom strengbrud, kovalent bindingsbrud eller nukleotidbase- og sukkerskader kan føre til cellealdring, celledød eller mutationer og relaterede patologier såsom aldring, infertilitet, neurodegenerative sygdomme og kræft (2, 61, 210, 233, 280, 290).
De processer, der igangsættes i levende celler på grund af VGIC-dysfunktion i deres cellemembraner, hvilket resulterer i OS, genetisk skade og relaterede patologier, giver en forklaring på den overflod af biologiske og sundhedsmæssige effekter, der er gennemgået i introduktionen (afsnit 1.2). Desuden kan dysfunktionen af VGIC’er forårsaget af menneskeskabt EMF-eksponering og fører til OS også forklare EHS, da EHS er ledsaget af OS (197, 352), og faktisk skyldes det sandsynligvis kronisk OS. De patofysiologiske ændringer i CNS, der observeres ledsage af EHS [(se 190)] kan forklares ved, at neuroner har højere procentdele af VGIC’er, da VGIC’er (specifikt Na+ og K+ VGIC’er) er mediatorerne for transmissionen af nerveimpulserne (231, 271).
Flere studier har fundet, at ELF EMF’er inducerer epigenetiske ændringer i celler, hvilket almindeligvis resulterer i ændret genekspression. Sådanne ændringer omfatter methylering/demethylering af gener via aktivering/deaktivering af methyltransferaseenzymer, post-translationel modifikation af histonproteiner og ændring af mikroRNA-ekspression (353–355). Epigenetiske ændringer kan inducere betydelige ændringer i cellefunktionen og dermed en organismes sundhed. Da ROS påvirker cellesignalering (232) også relateret til epigenetiske ændringer (320, 354), kan de rapporterede epigenetiske effekter induceret af menneskeskabte ELF-EMF’er skyldes ROS-signalering, og den præsenterede mekanisme for EMF-induceret ROS (over)produktion giver en forklaring på dette. For eksempel kan EMF-induceret ROS interferere med DNA- eller histonmethyltransferaser og histondeacetylaser, hvilket resulterer i modifikationer af epigenomet i forskellige regioner, herunder promotorregionerne af tumorsuppressorgener, hvilket resulterer i deres hæmning/inaktivering og fører til kræftfremme (356, 357).
Ligesom vi gjorde, bemærkede Blank og Goodman (57, 58) også, at både ELF og “RF” (faktisk WC) EMF’er producerer lignende effekter, især ved at inducere syntese af stressproteiner i celler meget hurtigt (inden for et par minutter). For os er en tilsyneladende forklaring på de almindelige ELF- og RF/WC EMF-effekter, der undgik opmærksomhed, at “RF” EMF’er påvirker celler ikke ved deres bærerkomponenter (RF), men ved deres ELF-komponenter af pulsering og modulering. Som dette studie viser, er det faktisk kun ELF EMF’er, der inducerer de ikke-termiske biologiske effekter, og de virker ikke direkte på DNA, men indirekte gennem VGIC dysfunktion og deraf følgende induktion af OS (2, 210). Endvidere følger det, at rene RF EMF’er kun kan inducere opvarmning ved tilstrækkeligt høje intensiteter og frekvenser (3).
Som dokumenteret her kan menneskeskabte EMF’er på miljømæssigt eksisterende niveauer ionisere levende væv gennem effekten af den genererede ROS/OS. Det er gennem ROS’ effekten, at skaden findes i DNA’et efter menneskeskabte og især WC EMF-eksponeringer. Der er en tæt sammenhæng mellem menneskeskabte EMF-eksponeringer, VGIC-dysfunktion, OS og DNA-/cellulær skade.
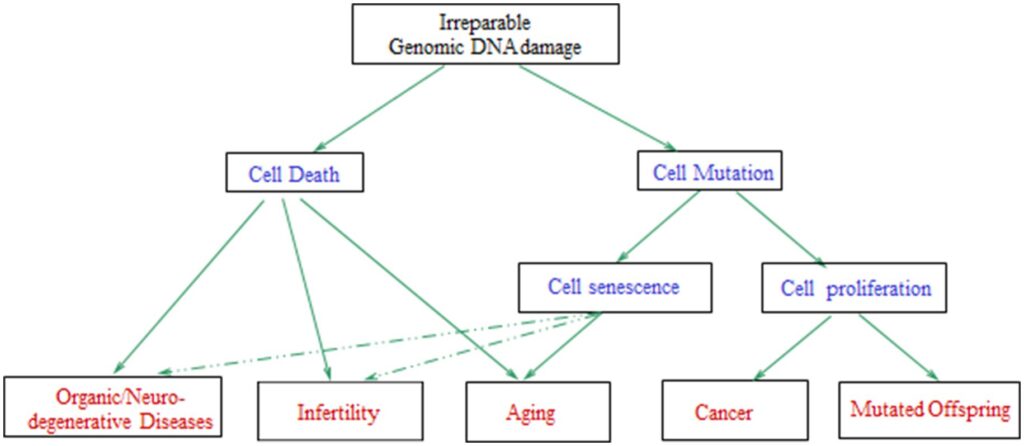
For celler med uopretteligt beskadiget genomisk DNA er mulige udfald cellealdring eller celledød (som kan resultere i aldring, organiske/neurodegenerative sygdomme og/eller reproduktive vanskeligheder), kræft eller muterede afkom (Figur 7), afhængigt af celletype, de specifikke biologiske/miljømæssige forhold og organismens tilstand (2, 210). Således forklarer DNA-skader induceret af OS de patologier, der er forbundet med kronisk eksponering for menneskeskabte EMF’er, såsom infertilitet og kræft.
Menneskeskabte EMF’er, og især de mest skadelige fra WC-antenner/-apparater og højspændingsledninger, er blevet en ny realitet i det moderne liv og udsætter milliarder af mennesker dagligt for eksponering (7, 50). Selvom de er betydeligt mindre cytotoksiske end radioaktivitet eller visse giftige kemikalier, repræsenterer de et evolutionært nyt og særdeles vedvarende dagligt cytotoksisk middel, mod hvilket eksisterende reparationsmekanismer muligvis ikke er effektive nok. Især hos personer, der allerede er genetisk eller epigenetisk kompromitteret.
Terapeutiske effekter af menneskeskabte EMF’er er også blevet rapporteret i litteraturen, især af pulserende ELF EMF’er og specifikt i knoglebrudsheling (238, 358–363). Ændrede intracellulære calciumniveauer er også blevet rapporteret at ledsage sådanne effekter, og den samme biofysiske mekanisme for induceret VGIC gating ser ud til at være involveret i både de skadelige og terapeutiske effekter af menneskeskabte EMF’er (238, 359, 363). Flere forfattere taler om terapeutiske effekter af pulserende ELF-EMF’er uden at specificere eller diskutere, hvilken parameter for EMF-eksponeringen der kan være den terapeutiske (364–366). Det kan føre til det fejlagtige indtryk, at enhver EMF med ELF/ULF-pulseringer kan være terapeutisk, hvilket naturligvis ikke er tilfældet, da f.eks. alle WC EMF’er består af sådanne pulseringer, og alligevel er de mest skadelige. Andre forfattere foreslår, at der er specifikke “gavnlige” eller “skadelige” frekvenser i ELF-området (367) uden at tage hensyn til IFO-VGIC-mekanismen, der er offentliggjort i næsten 25 år (og allerede refereret til i mere end tusind andre publikationer), som tydeligt viser ifølge almindeligt accepteret matematik, fysik og biologi, at bioaktiviteten af polariserede, sammenhængende og oscillerende EMF’er er proportional med feltintensiteten og omvendt proportional med feltfrekvensen, hvilket gør alle ULF/ELF-frekvenser meget bioaktive snarere end kun nogle specifikke (2, 5, 297, 305, 306). Grundlaget for EMF-bioaktivitet er således ikke nogle specifikke frekvenser, men polarisering og kohærens kombineret med lavfrekvent variabilitet (ved enhver ULF/ELF-frekvens), hvor jo lavere frekvens, jo mere bioaktivt er feltet (Ligning 16). Når en EMF er polariseret, sammenhængende og langsomt varierende, kan vi ikke udelukke muligheden for, at resonansfænomener finder sted ved specifikke fysiologiske ULF/ELF-frekvenser. Vi ville imidlertid ikke forvente, at sådanne fænomener, hvis de opstår, ville være særligt intense, navnlig under faktiske dæmpningsforhold i celler og ionkanaler (368).
Efter vores mening er en betingelse for, at en anvendt EMF har en terapeutisk virkning, at den simulere naturlige EMF’er eller fysiologiske endogene cellulære signaler. Når vi ved, at de mest bioaktive polariserede og sammenhængende EMF’er er ULF/ELF-dem, er det kritiske spørgsmål for en anvendt ULF/ELF EMF, om dens inkluderede frekvenser (og andre parametre såsom bølgeform, polaritet osv.) forstærker eller annullerer den endogene fysiologiske elektriske aktivitet af cellerne, som er ansvarlig for den specifikke terapeutiske virkning (49, 210, 369). Grundfrekvensen af de naturlige atmosfæriske “Schumann” elektromagnetiske resonanser (7,83 Hz) og dens harmoni er detekteret i menneskets/dyrets hjerneaktivitet, og de fysiske parametre for elektromagnetisk hjerneaktivitet og atmosfærisk lyn viser bemærkelsesværdige ligheder (369–371). Derfor har vi foreslået (210), at de terapeutiske effekter af pulserende elektromagnetiske felter forventes at være optimale ved pulserende frekvenser, der falder sammen med Schumann-frekvenserne, eller de endogene ioniske svingninger i celler (49). Faktisk fandt Yan et al. (372), at pulser ved en ELF-gentagelseshastighed, der falder sammen med den grundlæggende Schumann-frekvens 7,83 Hz, hæmmer proliferation og inducerer apoptose af kræftceller, mens dette ikke forekommer med normale celler. Det skal verificeres yderligere, og der er bestemt væsentlige begrænsninger: Alle menneskeskabte EMF’er er fuldt polariserede og sammenhængende, noget der ikke forekommer med de naturlige EMF’er, som kun er delvist polariserede ved visse lejligheder (5). Det ser ud til at være grunden til, at langt de fleste effekter af menneskeskabte EMF’er er skadelige, mens langt de fleste naturlige EMF’er kan være gavnlige.
Afslutningsvis giver IFO-VGIC-mekanismen, der forklarer VGIC-dysfunktion, og det efterfølgende OS, en omfattende biofysisk/biokemisk mekanisme, der forklarer overfloden af eksperimentelle og epidemiologiske resultater, der forbinder menneskeskabte EMF-eksponeringer med OS, DNA/cellulær skade og relaterede patologier såsom dårligt helbred, EHS, infertilitet, organiske/neurodegenerative sygdomme, kræft osv. Selvom de mekanistiske detaljer om, hvordan de ioniske forstyrrelser præcist stimulerer ROS-produktionen af deres kilder, skal udforskes yderligere, er det grundlæggende skema for den komplette EMF-bioeffektmekanisme allerede afsløret. De længe eksisterende eksperimentelle og epidemiologiske resultater, der forbinder eksponering for menneskeskabte EMF’er og DNA-skader, infertilitet og kræft, forklares nu af den omfattende mekanisme, der præsenteres her. Vi håber, at dette giver en bedre forståelse af den involverede forskning, et grundlag for fremtidig forskning samt etablering af biologisk relevante retningslinjer for EMF-eksponering for effektiv beskyttelse af folkesundheden og miljøet.
Interessekonflikt
Forfatterne erklærer, at forskningen blev udført i fravær af kommercielle eller finansielle forbindelser, der kunne fortolkes som en potentiel interessekonflikt.
Generativ AI-erklæring
Forfatterne erklærer, at der ikke blev brugt nogen Gen AI i skabelsen af dette manuskript.
Referencer
1. Misek, J, Jakus, J, Hamza Sladicekova, K, Zastko, L, Veternik, M, Jakusova, V, et al. Extremely low frequency magnetic fields emitted by cell phones. Front Phys. (2023) 11:1094921. doi: 10.3389/fphy.2023.1094921 Crossref Full Text | Google Scholar
2. Panagopoulos, DJ, Karabarbounis, A, Yakymenko, I, and Chrousos, GP. Human-made electromagnetic fields: ion forced-oscillation and voltage-gated ion channel dysfunction, oxidative stress and DNA damage. Int J Oncol. (2021) 59:92. doi: 10.3892/ijo.2021.5272
PubMed Abstract | Crossref Full Text | Google Scholar
3. Panagopoulos, DJ, Karabarbounis, A, and Lioliousis, C. Defining wireless communication (WC) electromagnetic fields (EMFs): A. Polarization is a principal property of all man-made EMFs; B. Modulation, pulsation, and variability are inherent parameters of WC EMFs; C. Most man-made EMF exposures are non-thermal; D. Measuring incident EMFs is more relevant than specific absorption rate (SAR); E. All man-made EMFs emit continuous waves, not photons; F. Differences from natural EMFs. Interaction with matter In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: Biological and health effects. Boca Raton: CRC Press (2022) Google Scholar
4. Pedersen, GF. Amplitude modulated RF fields stemming from a GSM/DCS-1800 phone. Wirel Netw. (1997) 3:489–98. doi: 10.1023/A:1019158712657 Crossref Full Text | Google Scholar
5. Panagopoulos, DJ, Johansson, O, and Carlo, GL. Polarization: a key difference between man-made and natural electromagnetic fields, in regard to biological activity. Sci Rep. (2015) 5:14914.
doi: 10.1038/srep14914 PubMed Abstract | Crossref Full Text | Google Scholar
6. Dubrov, AP. The geomagnetic field and life. New York: Plenum Press (1978). Google Scholar
7. Panagopoulos, DJ. Comparing DNA damage induced by Mobile telephony and other types of man-made electromagnetic fields. Mutation Res Rev. (2019) 781:53–62. doi: 10.1016/j.mrrev.2019.03.003
PubMed Abstract | Crossref Full Text | Google Scholar
8. Bawin, SM, Kaczmarek, LK, and Adey, WR. Effects of modulated VHF fields, on the central nervous system. Ann NY Acad Sci. (1975) 247:74–81. Google Scholar
9. Bawin, SM, Adey, WR, and Sabbot, IM. Ionic factors in release of 45Ca2+ from chick cerebral tissue by electromagnetic fields. Proc Natl Acad Sci USA. (1978) 75:6314–8. Google Scholar
10. Blackman, CF, Benane, SG, Elder, JA, House, DE, Lampe, JA, and Faulk, JM. Induction of calcium-ion efflux from brain tissue by radiofrequency radiation: effect of sample number and modulation frequency on the power-density window. Bioelectromagnetics. (1980) 1:35–43. doi: 10.1002/bem.2250010104
PubMed Abstract | Crossref Full Text | Google Scholar
11. Blackman, CF, Benane, SG, Kinney, LS, Joines, WT, and House, DE. Effects of ELF fields on calcium-ion efflux from brain tissue in vitro. Radiat Res. (1982) 92:510–20.
PubMed Abstract | Google Scholar
12. Bolshakov, MA, and Alekseev, SI. Bursting responses of Lymnea neurons to microwave radiation. Bioelectromagnetics. (1992) 13:119–29. doi: 10.1002/bem.2250130206
PubMed Abstract | Crossref Full Text | Google Scholar
13. Byus, CV, Lundak, RL, Fletcher, RM, and Adey, WR. Alterations in protein kinase activity following exposure of cultured lymphocytes to modulated microwave fields. Bioelectromagnetics. (1984) 5:341–51. doi: 10.1002/bem.2250050307
PubMed Abstract | Crossref Full Text | Google Scholar
14. Byus, CV, Kartum, K, Pieper, SE, and Adey, WR. Ornithine decarboxylase activity in liver cells is enhanced by low-level amplitude modulated microwave fields. Cancer Res. (1988) 48:4222–6.
PubMed Abstract | Google Scholar
15. Campisi, A, Gulino, M, Acquaviva, R, Bellia, P, Raciti, G, Grasso, R, et al. Reactive oxygen species levels and DNA fragmentation on astrocytes in primary culture after acute exposure to low intensity microwave electromagnetic field. Neurosci Lett. (2010) 473:52–5. doi: 10.1016/j.neulet.2010.02.018
Crossref Full Text | Google Scholar
16. Franzellitti, S, Valbonesi, P, Ciancaglini, N, Biondi, C, Contin, A, Bersani, F, et al. Transient DNA damage induced by high-frequency electromagnetic fields (GSM 1.8 GHz) in the human trophoblast HTR-8/SVneo cell line evaluated with the alkaline comet assay. Mutat Res. (2010) 683:35–42.
doi: 10.1016/j.mrfmmm.2009.10.004
PubMed Abstract | Crossref Full Text | Google Scholar
17. Frei, M, Jauchem, J, and Heinmets, F. Physiological effects of 2.8 GHz radio-frequency radiation: a comparison of pulsed and continuous-wave radiation. J Microw Power Electromagn Energy. (1988) 23:2.
Google Scholar
18. Goodman, EM, Greenebaum, B, and Marron, MT. Effects of electro- magnetic fields on molecules and cells. Int Rev Cytologia. (1995) 158:279–338. Google Scholar
19. Hinrikus, H, Bachmann, M, Lass, J, Tomson, R, and Tuulik, V. Effect of 7, 14 and 21 Hz modulated 450 MHz microwave radiation on human electroencephalographic rhythms. Int J Radiat Biol. (2008) 84:69–79. doi: 10.1080/09553000701691679 PubMed Abstract | Crossref Full Text | Google Scholar
20. Höytö, A, Luukkonen, J, Juutilainen, J, and Naarala, J. Proliferation, oxidative stress and cell death in cells exposed to 872 MHz radiofrequency radiation and oxidants. Radiat Res. (2008) 170:235–43. doi: 10.1667/RR1322.1 PubMed Abstract | Crossref Full Text | Google Scholar
21. Huber, R, Treyer, V, Borbely, AA, Schuderer, J, Gottselig, JM, Landolt, HP, et al. Electromagnetic fields, such as those from mobile phones, alter regional cerebral blood flow and sleep and waking EEG. J Sleep Res. (2002) 11:289–95. doi: 10.1046/j.1365-2869.2002.00314.x
PubMed Abstract | Crossref Full Text | Google Scholar
22. Lin-Liu, S, and Adey, WR. Low frequency amplitude modulated microwave fields change calcium efflux rates from synaptosomes. Bioelectromagnetics. (1982) 3:309–22. doi: 10.1002/bem.2250030303
PubMed Abstract | Crossref Full Text | Google Scholar
23. Litovitz, TA, Krause, D, Penafiel, M, Elson, EC, and Mullins, JM. The role of coherence time in the effect of microwaves on ornithine decarboxylase activity. Bioelectromagnetics. (1993) 14:395–403. doi: 10.1002/bem.2250140502
PubMed Abstract | Crossref Full Text | Google Scholar
24. Mohammed, HS, Fahmy, HM, Radwan, NM, and Elsayed, AA. Non-thermal continuous and modulated electromagnetic radiation fields effects on sleep EEG of rats. J Adv Res. (2013) 4:181–7. doi: 10.1016/j.jare.2012.05.005
PubMed Abstract | Crossref Full Text | Google Scholar
25. Penafiel, LM, Litovitz, T, Krause, D, Desta, A, and Mullins, MJ. Role of modulation on the effect of microwaves on ornithine decarboxylase activity in L929 cells. Bioelectromagnetics. (1997) 18:132–41. doi: 10.1002/(SICI)1521-186X(1997)18:2<132::AID-BEM6>3.0.CO;2-3
PubMed Abstract | Crossref Full Text | Google Scholar
26. Somosy, Z, Thuroczy, G, Kubasova, T, Kovacs, J, and Szabo, LD. Effects of modulated and continuous microwave irradiation on the morphology and cell surface negative charge of 3T3 fibroblasts. Scanning Microsc. (1991) 5:1145–55. Google Scholar
27. Thuroczy, G, Kubinyi, G, Bodo, M, Bakos, J, and Szabo, LD. Simultaneous response of brain electrical activity (EEG) and cerebral circulation (REG) to microwave exposure in rats. Rev Environ Health. (1994) 10:135–48. doi: 10.1515/REVEH.1994.10.2.135
PubMed Abstract | Crossref Full Text | Google Scholar
28. Van der Meer, JN, Eisma, YB, Meester, R, Jacobs, M, and Nederveen, AJ. Effects of mobile phone electromagnetic fields on brain waves in healthy volunteers. Sci Rep. (2023) 13:21758. doi: 10.1038/s41598-023-48561-z
PubMed Abstract | Crossref Full Text | Google Scholar
29. Veyret, B, Bouthet, C, Deschaux, P, de Seze, R, Geffard, M, Joussot-Dubien, J, et al. Antibody responses of mice exposed to low-power microwaves under combined, pulse-and-amplitude modulation. Bioelectromagnetics. (1991) 12:47–56. doi: 10.1002/bem.2250120107
PubMed Abstract | Crossref Full Text | Google Scholar
30. Mohammed, HS. Effects of wireless communication electromagnetic fields on human and animal brain activity In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press (2022) Google Scholar
31. Aydin, M, Turk, G, Yuksel, M, Cevik, A, Apaydin, A, and Yilmaz, S. Effect of electromagnetic field on the sperm characteristics and histopathological status of testis in rats. Med Weter. (2007) 63:178–83.
Google Scholar
32. Azanza, MJ, Perez Bruzon, RN, Lederer, D, et al. Reversibility of the effects induced on the spontaneous bioelectric activity of neurons under exposure to 8.3 and 217.0 Hz low intensity magnetic fields. 2nd Int. Workshop Biol Effects of EMFs, Rhodes, Greece. (2002). p. 651–659.
Google Scholar
33. Hong, R, Zhang, Y, Liu, Y, and Weng, EQ. Effects of extremely low frequency electromagnetic fields on DNA of testicular cells and sperm chromatin structure in mice. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi. (2005) 23:414–7. PubMed Abstract | Google Scholar
34. Ivancsits, S, Diem, E, Pilger, A, Rüdiger, HW, and Jahn, O. Induction of DNA strand breaks by intermittent exposure to extremely-low-frequency electromagnetic fields in human diploid fibroblasts. Mutat Res. (2002) 519:1–13. doi: 10.1016/S1383-5718(02)00109-2
PubMed Abstract | Crossref Full Text | Google Scholar
35. Ivancsits, S, Diem, E, Jahn, O, and Rüdiger, HW. Intermittent extremely low frequency electromagnetic fields cause DNA damage in a dose-dependent way. Int Arch Occup Environ Health. (2003) 76:431–6.
doi: 10.1007/s00420-003-0446-5
PubMed Abstract | Crossref Full Text | Google Scholar
36. Lee, JS, Ahn, SS, Jung, KC, Kim, YW, and Lee, SK. Effects of 60 Hz electromagnetic field exposure on testicular germ cell apoptosis in mice. Asian J Androl. (2004) 6:29–34. Google Scholar
37. Lee, SK, Park, S, Gimm, YM, and Kim, YW. Extremely low frequency magnetic fields induce spermatogenic germ cell apoptosis: possible mechanism. Biomed Res Int. (2014) 2014:567183:1–8.
doi: 10.1155/2014/567183 PubMed Abstract | Crossref Full Text | Google Scholar
38. Ma, TH, and Chu, KC. Effect of the extremely low frequency (ELF) electromagnetic field (EMF) on developing embryos of the fruit fly (Drosophila melanogaster L.). Mutat Res. (1993) 303:35–9.
doi: 10.1016/0165-7992(93)90006-H
PubMed Abstract | Crossref Full Text | Google Scholar
39. Moghadam, MK, Firoozabadi, SM, and Janahmadi, M. 50 Hz alternating extremely low frequency magnetic fields affect excitability, firing and action potential shape through interaction with ionic channels in snail neurones. Environmentalist. (2008) 28:341–7. doi: 10.1007/s10669-007-9143-3
Crossref Full Text | Google Scholar
40. Moghadam, MK, Firoozabadi, SM, and Janahmadi, M. Reduction of F1 neuronal excitability by exposure to 217 Hz magnetic fields from GSM 900 mobile phone. Cell J. (2009) 11:176–83.
Google Scholar
41. Moghadam, MK, Firoozabadi, M, and Janahmadi, M. Effects of weak environmental magnetic fields on the spontaneous bioelectrical activity of snail neurons. J Membr Biol. (2011) 240:63–71.
doi: 10.1007/s00232-011-9344-z
PubMed Abstract | Crossref Full Text | Google Scholar
42. Panagopoulos, DJ, Karabarbounis, A, and Lioliousis, C. ELF alternating magnetic field decreases reproduction by DNA damage induction. Cell Biochem Biophys. (2013) 67:703–16.
doi: 10.1007/s12013-013-9560-5
PubMed Abstract | Crossref Full Text | Google Scholar
43. Santini, MT, Ferrante, A, Rainaldi, G, Indovina, P, and Indovina, PL. Extremely low frequency (ELF) magnetic fields and apoptosis: a review. Int J Radiat Biol. (2005) 81:1–11.
doi: 10.1080/09553000400029502
PubMed Abstract | Crossref Full Text | Google Scholar
44. Walleczek, J. Electromagnetic field effects on cells of the immune system: the role of calcium signaling. FASEB J. (1992) 6:3177–85. doi: 10.1096/fasebj.6.13.1397839
PubMed Abstract | Crossref Full Text | Google Scholar
45. Górski, R, Kotwicka, M, Skibińska, I, Jendraszak, M, and Wosiński, S. Effect of low-frequency electric field screening on motility of human sperm. Ann Agric Environ Med. (2020) 27:427–34.
doi: 10.26444/aaem/116019
PubMed Abstract | Crossref Full Text | Google Scholar
46. Karbalay-Doust, S, Darabyan, M, Sisakht, M, Haddadi, G, Sotoudeh, N, Haghani, M, et al. Extremely low frequency-electromagnetic fields (ELF-EMF) can decrease spermatocyte count and motility and change testicular tissue. J Biomed Phys Eng. (2023) 13:135–46. doi: 10.31661/jbpe.v0i0.2011-1234
PubMed Abstract | Crossref Full Text | Google Scholar
47. Mahna, A, Firoozabadi, SM, and Atashi, A. Cell phone and breast cancer: the cell phone-generated pulsed 217Hz ELF magnetic field increases angiogenesis. Iran J Med Phys. (2021) 18:421–9.
doi: 10.22038/IJMP.2020.52303.1859 Crossref Full Text | Google Scholar
48. Mansourian, M, Firoozabadi, M, and Hassan, ZM. The role of 217-Hz ELF magnetic fields emitted from GSM mobile phones on electrochemotherapy mechanisms. Electromagn Biol Med. (2020) 39:239–49.
doi: 10.1080/15368378.2020.1762635
PubMed Abstract | Crossref Full Text | Google Scholar
49. Panagopoulos, DJ. Electromagnetic interaction between environmental fields and living systems determines health and well-being In: Electromagnetic fields: principles, engineering applications and biophysical effects. K Myung-Hee and Y Sang-Ook (editors). New York, USA: Nova Science Publishers (2013) Google Scholar
50. Panagopoulos, DJ. Electromagnetic fields of wireless communications: biological and health effects. 1st ed. Boca Raton: CRC Press (2023). Google Scholar
51. Persinger, MA. ELF and VLF electromagnetic fields. New York: Plenum Press (1974). Google Scholar
52. Wever, R. The circadian system of man: results of experiments under temporal isolation. New York: Springer-Verlag (1979). Google Scholar
53. Panagopoulos, DJ. Man-made electromagnetic radiation is not quantized In: A Reimer, editor. Horizons in world physics, vol. 296. New York: Nova Science Publishers (2018). 1–57.
Google Scholar
54. Puranen, L, and Jokela, K. Radiation hazard assessment of pulsed microwave radars. J Microw Power Electromagn Energy. (1996) 31:165–77. doi: 10.1080/08327823.1996.11688307
PubMed Abstract | Crossref Full Text | Google Scholar
55. Pirard, W, and Vatovez, B. Study of pulsed character of radiation emitted by wireless telecommunication systems. Liège, Belgium: Institut scientifique de service public.
(Available in: https://www.issep.be/wp-content/uploads/7IWSBEEMF_B-Vatovez_W-Pirard.pdf)
Google Scholar
56. Panagopoulos, DJ. Comments on Pall’s “millimeter (MM) wave and microwave frequency radiation produce deeply penetrating effects: the biology and the physics”. Rev Environ Health. (2021) 37:295–7. doi: 10.1515/reveh-2021-0090
PubMed Abstract | Crossref Full Text | Google Scholar
57. Blank, M, and Goodman, R. Electromagnetic fields stress living cells. Pathophysiology. (2009) 16:71–8. doi: 10.1016/j.pathophys.2009.01.006
PubMed Abstract | Crossref Full Text | Google Scholar
58. Blank, M, and Goodman, R. DNA is a fractal antenna in electromagnetic fields. Int J Radiat Biol. (2011) 87:409–15. doi: 10.3109/09553002.2011.538130
PubMed Abstract | Crossref Full Text | Google Scholar
59. Henshaw, DL, and Philips, A. A mechanistic understanding of human magnetoreception validates the phenomenon of electromagnetic hypersensitivity (EHS). Int J Radiat Biol. (2024):1–19. Google Scholar
60. Roux, D, Vian, A, Girard, S, Bonnet, P, Paladian, F, Davies, E, et al. Electromagnetic fields (900MHz) evoke consistent molecular responses in tomato plants. Physiol Plant. (2006) 128:283–8.
doi: 10.1111/j.1399-3054.2006.00740.x Crossref Full Text | Google Scholar
61. Phillips, JL, Singh, NP, and Lai, H. Electromagnetic fields and DNA damage. Pathophysiology. (2009) 16:79–88. doi: 10.1016/j.pathophys.2008.11.005
PubMed Abstract | Crossref Full Text | Google Scholar
62. Yakymenko, I, Tsybulin, O, Sidorik, E, Henshel, D, Kyrylenko, O, and Kyrylenko, S. Oxidative mechanisms of biological activity of low-intensity radiofrequency radiation. Electromagn Biol Med. (2016) 35:186–202. doi: 10.3109/15368378.2015.1043557
PubMed Abstract | Crossref Full Text | Google Scholar
63. Yakymenko, I, and Tsybulin, O. Oxidative stress induced by wireless communication electromagnetic fields In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press (2022) Google Scholar
64. Jagetia, GC. Genotoxic effects of wireless communication electromagnetic fields In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press, Taylor and Francis (2022) Google Scholar
65. Miller, K, Harrison, K, Martin, JH, Nixon, B, and De Iuliis, GN. The impacts of wireless communication electromagnetic fields on human reproductive biology In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC press, Taylor and Francis (2022)
Google Scholar
66. Panagopoulos, DJ. DNA and chromosome damage in human and animal cells induced by mobile telephony electromagnetic fields and other stressors In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press (2022)
Google Scholar
67. Agarwal, A, Desai, NR, Makker, K, Varghese, A, Mouradi, R, Sabanegh, E, et al. Effects of radiofrequency electromagnetic waves (RF-EMW) from cellular phones on human ejaculated semen: an in vitro pilot study. Fertil Steril. (2009) 92:1318–25. doi: 10.1016/j.fertnstert.2008.08.022
PubMed Abstract | Crossref Full Text | Google Scholar
68. Aitken, RJ, Bennetts, LE, Sawyer, D, Wiklendt, AM, and King, BV. Impact of radio frequency electromagnetic radiation on DNA integrity in the male germline. Int J Androl. (2005) 28:171–9. doi: 10.1111/j.1365-2605.2005.00531.x
PubMed Abstract | Crossref Full Text | Google Scholar
69. Banerjee, S, Singh, NN, Sreedhar, G, and Mukherjee, S. Analysis of the genotoxic effects of Mobile phone radiation using buccal micronucleus assay: a comparative evaluation. J Clin Diagn Res. (2016) 10:ZC82–5. doi: 10.7860/JCDR/2016/17592.7505
PubMed Abstract | Crossref Full Text | Google Scholar
70. Belyaev, IY, Hillert, L, Protopopova, M, Tamm, C, Malmgren, LOG, Persson, BRR, et al. 915 MHz microwaves and 50 Hz magnetic field affect chromatin conformation and 53BP1 foci in human lymphocytes from hypersensitive and healthy persons. Bioelectromagnetics. (2005) 26:173–84.
doi: 10.1002/bem.20103
PubMed Abstract | Crossref Full Text | Google Scholar
71. Burlaka, A, Tsybulin, O, Sidorik, E, Lukin, S, Polishuk, V, Tsehmistrenko, S, et al. Overproduction of free radical species in embryonic cells exposed to low intensity radiofrequency radiation. Exp Oncol. (2013) 35:219–25. PubMed Abstract | Google Scholar
72. Chavdoula, ED, Panagopoulos, DJ, and Margaritis, LH. Comparison of biological effects between continuous and intermittent exposure to GSM-900 MHz mobile phone radiation. Detection of apoptotic cell death features. Mutation Res. (2010) 700:51–61. doi: 10.1016/j.mrgentox.2010.05.008
PubMed Abstract | Crossref Full Text | Google Scholar
73. Daroit, NB, Visioli, F, Magnusson, AS, Vieira, GR, and Rados, PV. Cell phone radiation effects on cytogenetic abnormalities of oral mucosal cells. Braz Oral Res. (2015) 29:1–8.
doi: 10.1590/1807-3107BOR-2015.vol29.0114
PubMed Abstract | Crossref Full Text | Google Scholar
74. De Iuliis, GN, Newey, RJ, King, BV, and Aitken, RJ. Mobile phone radiation induces reactive oxygen species production and DNA damage in human spermatozoa in vitro. PLoS One. (2009) 4:e6446.
doi: 10.1371/journal.pone.0006446
PubMed Abstract | Crossref Full Text | Google Scholar
75. De Iuliis, GN, Thomson, LK, Mitchell, LA, Finnie, JM, Koppers, AJ, et al. DNA damage in human spermatozoa is highly correlated with the efficiency of chromatin remodeling and the formation of 8-hydroxy-2′-deoxyguanosine, a marker of oxidative stress. Biol Reprod. (2009) 81:517–24.
doi: 10.1095/biolreprod.109.076836
PubMed Abstract | Crossref Full Text | Google Scholar
76. Delgado, JMR. Biological effects of extremely low frequency electromagnetic fields. J Bioelectr. (1985) 4:75–92. doi: 10.3109/15368378509040362 Crossref Full Text | Google Scholar
77. Diem, E, Schwarz, C, Adlkofer, F, Jahn, O, and Rudiger, H. Non-thermal DNA breakage by mobile-phone radiation (1800 MHz) in human fibroblasts and in transformed GFSH-R17 rat granulosa cells in vitro. Mutat Res. (2005) 583:178–83. doi: 10.1016/j.mrgentox.2005.03.006
PubMed Abstract | Crossref Full Text | Google Scholar
78. Ferreira, AR, Knakievicz, T, de Bittencourt Pasquali, MA, Gelain, DP, Dal-Pizzol, F, Fernández, CER, et al. Ultra high frequency-electromagnetic field irradiation during pregnancy leads to an increase in erythrocytes micronuclei incidence in rat offspring. Life Sci. (2006) 80:43–8050.
doi: 10.1016/j.lfs.2006.08.018 Crossref Full Text | Google Scholar
79. Garaj-Vrhovac, V, Horvat, D, and Koren, Z. The effect of microwave radiation on the cell genome. Mutat Res. (1990) 243:87–93. doi: 10.1016/0165-7992(90)90028-I
PubMed Abstract | Crossref Full Text | Google Scholar
80. Garaj-Vrhovac, V, Horvat, D, and Koren, Z. The relationship between colony-forming ability, chromosome aberrations and incidence of micronuclei in V79 Chinese hamster cells exposed to microwave radiation. Mutat Res. (1991) 263:143–9. doi: 10.1016/0165-7992(91)90054-8
PubMed Abstract | Crossref Full Text | Google Scholar
81. Garaj-Vrhovac, V, Fucić, A, and Horvat, D. The correlation between the frequency of micronuclei and specific chromosome aberrations in human lymphocytes exposed to microwave radiation in vitro. Mutat Res. (1992) 281:181–6. doi: 10.1016/0165-7992(92)90006-4
PubMed Abstract | Crossref Full Text | Google Scholar
82. Guler, G, Tomruk, A, Ozgur, E, and Seyhan, N. The effect of radiofrequency radiation on DNA and lipid damage in non-pregnant and pregnant rabbits and their newborns. Gen Physiol Biophys. (2010) 29:59–60. doi: 10.4149/gpb_2010_01_59
PubMed Abstract | Crossref Full Text | Google Scholar
83. Ji, S, Oh, E, Sul, D, Choi, JW, Park, H, and Lee, E. DNA damage of lymphocytes in volunteers after 4 hours use of Mobile phone. J Prev Med Public Health. (2004) 37:373–80. Google Scholar
84. Koana, T, Okada, MO, Takashima, Y, Ikehata, M, and Miyakoshi, J. Involvement of eddy currents in the mutagenicity of ELF magnetic fields. Mutat Res. (2001) 476:55–62. doi: 10.1016/S0027-5107(01)00082-3
PubMed Abstract | Crossref Full Text | Google Scholar
85. Lai, H, and Singh, NP. Acute low-intensity microwave exposure increases DNA single-strand breaks in rat brain cells. Bioelectromagnetics. (1995) 16:207–10. doi: 10.1002/bem.2250160309
PubMed Abstract | Crossref Full Text | Google Scholar
86. Lai, H, and Singh, NP. Single- and double-strand DNA breaks in rat brain cells after acute exposure to radiofrequency electromagnetic radiation. Int J Radiat Biol. (1996) 69:513–21.
doi: 10.1080/095530096145814
PubMed Abstract | Crossref Full Text | Google Scholar
87. Lai, H, and Singh, NP. Acute exposure to a 60 Hz magnetic field increases DNA strand breaks in rat brain cells. Bioelectromagnetics. (1997) 18:156–65.
doi: 10.1002/(SICI)1521-186X(1997)18:2<156::AID-BEM8>3.0.CO;2-1
PubMed Abstract | Crossref Full Text | Google Scholar
88. Lee, KS, Choi, JS, Hong, SY, Son, TH, and Yu, K. Mobile phone electromagnetic radiation activates MAPK signaling and regulates viability in Drosophila. Bioelectromagnetics. (2008) 29:371–9.
doi: 10.1002/bem.20395
PubMed Abstract | Crossref Full Text | Google Scholar
89. Liu, C, Gao, P, Xu, SC, Wang, Y, Chen, CH, He, MD, et al. Mobile phone radiation induces mode-dependent DNA damage in a mouse spermatocyte-derived cell line: a protective role of melatonin. Int J Radiat Biol. (2013) 89:993–1001. doi: 10.3109/09553002.2013.811309
PubMed Abstract | Crossref Full Text | Google Scholar
90. Lixia, S, Yao, K, Kaijun, W, Deqiang, L, Huajun, H, Xiangwei, G, et al. Effects of 1.8 GHz radiofrequency field on DNA damage and expression of heat shock protein 70 in human lens epithelial cells. Mutat Res. (2006) 602:135–42. doi: 10.1016/j.mrfmmm.2006.08.010
PubMed Abstract | Crossref Full Text | Google Scholar
91. Luukkonen, J, Hakulinen, P, Mäki-Paakkanen, J, Juutilainen, J, and Naarala, J. Enhancement of chemically induced reactive oxygen species production and DNA damage in human SH-SY5Y neuroblastoma cells by 872 MHz radiofrequency radiation. Mutat Res. (2009) 662:54–8.
doi: 10.1016/j.mrfmmm.2008.12.005
PubMed Abstract | Crossref Full Text | Google Scholar
92. Mailankot, M, Kunnath, AP, Jayalekshmi, H, Koduru, B, and Valsalan, R. Radio frequency electromagnetic radiation (RF-EMR) from GSM (0.9/1.8GHz) mobile phones induces oxidative stress and reduces sperm motility in rats. Clinics. (2009) 64:561–5. doi: 10.1590/S1807-59322009000600011
PubMed Abstract | Crossref Full Text | Google Scholar
93. Markova, E, Hillert, L, Malmgren, L, Persson, BR, and Belyaev, IY. Microwaves from GSM mobile telephones affect 53BP1 and gamma-H2AX foci in human lymphocytes from hypersensitive and healthy persons. Environ Health Perspect. (2005) 113:1172–7. doi: 10.1289/ehp.7561
PubMed Abstract | Crossref Full Text | Google Scholar
94. Mausset-Bonnefont, AL, et al. Acute exposure to GSM 900-MHz electromagnetic fields induces glial reactivity and biochemical modifications in the rat brain. Neurobiol Dis. (2004) 17:445–54.
doi: 10.1016/j.nbd.2004.07.004
PubMed Abstract | Crossref Full Text | Google Scholar
95. Mihai, CT, Rotinberg, P, Brinza, F, and Vochita, G. Extremely low-frequency electromagneticfields cause DNA strand breaks in normal cells. J Environ Health Sci Eng. (2014) 12:15. doi: 10.1186/2052-336X-12-15
PubMed Abstract | Crossref Full Text | Google Scholar
96. Nikolova, T, Czyz, J, Rolletschek, A, Blyszczuk, P, Fuchs, J, Jovtchev, G, et al. Electromagnetic fields affect transcript levels of apoptosis-related genes in embryonic stem cell-derived neural progenitor cells. FASEB J. (2005) 19:1686–8. doi: 10.1096/fj.04-3549fje
PubMed Abstract | Crossref Full Text | Google Scholar
97. Panagopoulos, DJ, Chavdoula, ED, Nezis, IP, and Margaritis, LH. Cell death induced by GSM 900MHz and DCS 1800MHz Mobile telephony radiation. Mutat Res. (2007) 626:69–78.
doi: 10.1016/j.mrgentox.2006.08.008
PubMed Abstract | Crossref Full Text | Google Scholar
98. Panagopoulos, DJ, Chavdoula, ED, and Margaritis, LH. Bioeffects of Mobile telephony radiation in relation to its intensity or distance from the antenna. Int J Radiat Biol. (2010) 86:345–57.
doi: 10.3109/09553000903567961
PubMed Abstract | Crossref Full Text | Google Scholar
99. Panagopoulos, DJ. Effect of microwave exposure on the ovarian development of Drosophila melanogaster. Cell Biochem Biophys. (2012) 63:121–32. doi: 10.1007/s12013-012-9347-0
PubMed Abstract | Crossref Full Text | Google Scholar
100. Pesnya, DS, and Romanovsky, AV. Comparison of cytotoxic and genotoxic effects of plutonium-239 alpha particles and mobile phone GSM 900 radiation in the Allium cepa test. Mutat Res. (2013) 750:27–33. doi: 10.1016/j.mrgentox.2012.08.010
PubMed Abstract | Crossref Full Text | Google Scholar
101. Sarkar, S, Ali, S, and Behari, J. Effect of low power microwave on the mouse genome: a direct DNA analysis. Mutat Res. (1994) 320:141–7. doi: 10.1016/0165-1218(94)90066-3
PubMed Abstract | Crossref Full Text | Google Scholar
102. Sokolovic, D, et al. Melatonin reduces oxidative stress induced by chronic exposure of microwave radiation from Mobile phones in rat brain. J Radiat Res. (2008) 49:579–86. doi: 10.1269/jrr.07077
PubMed Abstract | Crossref Full Text | Google Scholar
103. Svedenstal, BM, Johanson, KJ, and Mild, KH. DNA damage induced in brain cells of CBA mice exposed to magnetic fields. In Vivo. (1999) 13:551–2.
PubMed Abstract | Google Scholar
104. Tomruk, A, Guler, G, and Dincel, AS. The influence of 1800 MHz GSM-like signals on hepatic oxidative DNA and lipid damage in nonpregnant, pregnant, and newly born rabbits. Cell Biochem Biophys. (2010) 56:39–47. doi: 10.1007/s12013-009-9068-1
PubMed Abstract | Crossref Full Text | Google Scholar
105. Winker, R, Ivancsits, S, Pilger, A, Adlkofer, F, and Rüdiger, HW. Chromosomal damage in human diploid fibroblasts by intermittent exposure to extremely low-frequency electromagnetic fields. Mutat Res. (2005) 585:43–9. doi: 10.1016/j.mrgentox.2005.04.013
PubMed Abstract | Crossref Full Text | Google Scholar
106. Yadav, AS, and Sharma, MK. Increased frequency of micronucleated exfoliated cells among humans exposed in vivo to mobile telephone radiations. Mutat Res. (2008) 650:175–80.
doi: 10.1016/j.mrgentox.2007.11.005
PubMed Abstract | Crossref Full Text | Google Scholar
107. Yan, JG, Agresti, M, Bruce, T, Yan, YH, Granlund, A, and Matloub, HS. Effects of cellular phone emissions on sperm motility in rats. Fertil Steril. (2007) 88:957–64. doi: 10.1016/j.fertnstert.2006.12.022
PubMed Abstract | Crossref Full Text | Google Scholar
108. Yao, K, et al. Electromagnetic noise inhibits radiofrequency radiation-induced DNA damage and reactive oxygen species increase in human lens epithelial cells. Mol Vis. (2008) 19:964–9.
Google Scholar
109. Zhang, DY, Xu, ZP, Chiang, H, Lu, DQ, and Zeng, QL. Effects of GSM 1800 MHz radiofrequency electromagnetic fields on DNA damage in Chinese hamster lung cells. Zhonghua Yu Fang Yi Xue Za Zhi. (2006) 40:149–52. PubMed Abstract | Google Scholar
110. Zhang, G, Yan, H, Chen, Q, Liu, K, Ling, X, Sun, L, et al. Effects of cell phone use on semen parameters: results from the MARHCS cohort study in Chongqing, China. Environ Int. (2016) 91:116–21.
doi: 10.1016/j.envint.2016.02.028
PubMed Abstract | Crossref Full Text | Google Scholar
111. D’Silva, MH, Swer, RT, Anbalagan, J, and Rajesh, B. Effect of radiofrequency radiation emitted from 2G and 3G cell phone on developing liver of Chick embryo – a comparative study. J Clin Diagn Res. (2017) 11:5–9. doi: 10.7860/JCDR/2017/26360.10275
PubMed Abstract | Crossref Full Text | Google Scholar
112. D’Silva, MH, Swer, RT, Anbalagan, J, and Bhargavan, R. Assessment of DNA damage in chick embryo brains exposed to 2G and 3G cell phone radiation using alkaline comet assay technique. J Clin Diagn Res. (2021) 15:AC01–4. doi: 10.7860/JCDR/2021/47115.14441
Crossref Full Text | Google Scholar
113. Gulati, S, Mosgoeller, W, Moldan, D, Kosik, P, Durdik, M, Jakl, L, et al. Evaluation of oxidative stress and genetic instability among residents near mobile phone base stations in Germany. Ecotoxicol Environ Saf. (2024) 279:116486. doi: 10.1016/j.ecoenv.2024.116486
PubMed Abstract | Crossref Full Text | Google Scholar
114. Gunes, M, Ates, K, Yalcin, B, Akkurt, S, Ozen, S, and Kaya, B. An evaluation of the genotoxic effects of electromagnetic radiation at 900 MHz, 1800 MHz, and 2100 MHz frequencies with a SMART assay in Drosophila melanogaster. Electromagn Biol Med. (2021) 40:254–63. doi: 10.1080/15368378.2021.1878210
Crossref Full Text | Google Scholar
115. Houston, BJ, Nixon, B, King, BV, Aitken, RJ, and De Iuliis, GN. Probing the origins of 1,800 MHz radio frequency electromagnetic radiation induced damage in mouse immortalized germ cells and spermatozoa in vitro. Front Public Health. (2018) 6:270. doi: 10.3389/fpubh.2018.00270
PubMed Abstract | Crossref Full Text | Google Scholar
116. Kesari, KK, Agarwal, A, and Henkel, R. Radiations and male fertility. Reprod Biol Endocrinol. (2018) 16:118. doi: 10.1186/s12958-018-0431-1
PubMed Abstract | Crossref Full Text | Google Scholar
117. Keskin, I, Karabulut, S, Kaplan, AA, Alagöz, M, Akdeniz, M, Tüfekci, KK, et al. Preliminary study on the impact of 900 MHz radiation on human sperm: an in vitro molecular approach. Reprod Toxicol. (2024) 130:108744. doi: 10.1016/j.reprotox.2024.108744
PubMed Abstract | Crossref Full Text | Google Scholar
118. Panagopoulos, DJ. Chromosome damage in human cells induced by UMTS Mobile telephony radiation. Gen Physiol Biophys. (2019) 38:445–54. doi: 10.4149/gpb_2019032
PubMed Abstract | Crossref Full Text | Google Scholar
119. Panagopoulos, DJ. Comparing chromosome damage induced by Mobile telephony radiation and a high caffeine dose. Effect of combination and exposure duration. Gen Physiol Biophys. (2020) 39:531–44. doi: 10.4149/gpb_2020036
Crossref Full Text | Google Scholar
120. Panagopoulos, DJ. Mobile telephony radiation exerts genotoxic action and significantly enhances the effects of gamma radiation in human cells. Gen Physiol Biophys. (2024) 43:103–20.
doi: 10.4149/gpb_2023036
PubMed Abstract | Crossref Full Text | Google Scholar
121. Shahin, S, Singh, SP, and Chaturvedi, CM. Mobile phone (1800MHz) radiation impairs female reproduction in mice, Mus musculus, through stress induced inhibition of ovarian and uterine activity. Reprod Toxicol. (2017) 73:41–60. doi: 10.1016/j.reprotox.2017.08.001
PubMed Abstract | Crossref Full Text | Google Scholar
122. Sharma, A, Shrivastava, S, and Shukla, S. Exposure of radiofrequency electromagnetic radiation on biochemical and pathological alterations. Neurol India. (2020) 68:1092–100.
doi: 10.4103/0028-3886.294554 Crossref Full Text | Google Scholar
123. Singh, KV, Prakash, C, Nirala, JP, Nanda, RK, and Rajamani, P. Acute radiofrequency electromagnetic radiation exposure impairs neurogenesis and causes neuronal DNA damage in the young rat brain. Neurotoxicology. (2023) 94:46–58. doi: 10.1016/j.neuro.2022.11.001
PubMed Abstract | Crossref Full Text | Google Scholar
124. Smith-Roe, SL, Wyde, ME, Stout, MD, Winters, JW, Hobbs, CA, Shepard, KG, et al. Evaluation of the genotoxicity of cell phone radiofrequency radiation in male and female rats and mice following subchronic exposure. Environ Mol Mutagen. (2019) 1:276–90. Google Scholar
125. Yakymenko, I, Burlaka, A, Tsybulin, I, Brieieva, I, Buchynska, L, Tsehmistrenko, S, et al. Oxidative and mutagenic effects of low intensity GSM 1800 MHz microwave radiation. Exp Oncol. (2018) 40:282–7.
doi: 10.31768/2312-8852.2018.40
PubMed Abstract | Crossref Full Text | Google Scholar
126. Yuan, LQ, Wang, C, Lu, DF, Zhao, XD, Tan, LH, and Chen, X. Induction of apoptosis and ferroptosis by a tumor suppressing magnetic field through ROS-mediated DNA damage. Aging. (2020) 12:3662–81.
doi: 10.18632/aging.102836 Crossref Full Text | Google Scholar
127. Cappucci, U, Casale, AM, Proietti, M, Marinelli, F, Giuliani, L, and Piacentini, L. WiFi related radiofrequency electromagnetic fields promote transposable element dysregulation and genomic instability in Drosophila melanogaster. Cells. (2022) 11:4036. doi: 10.3390/cells11244036
PubMed Abstract | Crossref Full Text | Google Scholar
128. Choi, J, Min, K, Jeon, S, Kim, N, Pack, JK, and Song, K. Continuous exposure to 1.7 GHz LTE electromagnetic fields increases intracellular reactive oxygen species to decrease human cell proliferation and induce senescence. Sci Rep. (2020) 10:9238. doi: 10.1038/s41598-020-65732-4
PubMed Abstract | Crossref Full Text | Google Scholar
129. Kim, JH, Jeon, S, Choi, HD, Lee, JH, Bae, JS, Kim, N, et al. Exposure to long-term evolution radiofrequency electromagnetic fields decreases neuroblastoma cell proliferation via Akt/mTOR-mediated cellular senescence. J Toxic Environ Health A. (2021) 84:846–57. doi: 10.1080/15287394.2021.1944944
PubMed Abstract | Crossref Full Text | Google Scholar
130. Shahin, S, Singh, VP, Shukla, RK, Dhawan, A, Gangwar, RK, Singh, SP, et al. 2.45 GHz microwave irradiation-induced oxidative stress affects implantation or pregnancy in mice, Mus musculus. Appl Biochem Biotechnol. (2013) 169:1727–51. doi: 10.1007/s12010-012-0079-9
PubMed Abstract | Crossref Full Text | Google Scholar
131. Czerwiński, M, Vian, A, Woodcock, BA, Goliński, P, Virto, LR, and Januszkiewicz, Ł. Do electromagnetic fields used in telecommunications affect wild plant species? A control impact study conducted in the field. Ecol Indic. (2023) 150:110267. doi: 10.1016/j.ecolind.2023.110267
Crossref Full Text | Google Scholar
132. Deena, K, Maadurshni, GB, Manivannan, J, and Sivasamy, R. Short-term exposure of 2.4 GHz electromagnetic radiation on cellular ROS generation and apoptosis in SH-SY5Y cell line and impact on developing chick embryo brain tissue. Mol Biol Rep. (2025) 52:144. doi: 10.1007/s11033-025-10217-8
PubMed Abstract | Crossref Full Text | Google Scholar
133. Özen, G, Kahvecioğlu, D, Bulut, İ, Erel, Ö, Neşelioğlu, S, Üstün, Y, et al. Effect of Mobile phone usage during pregnancy on total oxidant and antioxidant levels in cord blood. J Behcet Uz Children’s Hosp. (2023) 13:177–84. doi: 10.4274/jbuch.galenos.2023
Crossref Full Text | Google Scholar
134. Agarwal, A, Deepinder, F, Sharma, RK, Ranga, G, and Li, J. Effect of cell phone usage on semen analysis in men attending infertility clinic: an observational study. Fertil Steril. (2008) 89:124–8.
doi: 10.1016/j.fertnstert.2007.01.166
PubMed Abstract | Crossref Full Text | Google Scholar
135. Bakacak, M, Bostancı, MS, Attar, R, Yıldırım, ÖK, Yıldırım, G, Bakacak, Z, et al. The effects of electromagnetic fields on the number of ovarian primordial follicles: an experimental study. Kaohsiung J Med Sci. (2015) 31:287–92. doi: 10.1016/j.kjms.2015.03.004
PubMed Abstract | Crossref Full Text | Google Scholar
136. Gul, A, Celebi, H, and Uğraş, S. The effects of microwave emitted by cellular phones on ovarian follicles in rats. Arch Gynecol Obstet. (2009) 280:729–33. doi: 10.1007/s00404-009-0972-9
PubMed Abstract | Crossref Full Text | Google Scholar
137. Guzey, YZ, and Onal, AG. Effects of chronic exposure to 2G/3G cell phone radiation on in vitro maturation of bovine oocytes. Indian J Anim Res. (2018) 52:523–6. Google Scholar
138. Gye, MC, and Park, CJ. Effect of electromagnetic field exposure on the reproductive system. Clin Exp Reprod Med. (2012) 39:1–9. doi: 10.5653/cerm.2012.39.1.1
PubMed Abstract | Crossref Full Text | Google Scholar
139. La Vignera, S, Condorelli, RA, Vicardi, E, D’Agata, R, and Calogero, AE. Effects of the exposure to Mobile phones on male reproduction: a review of the literature. J Androl. (2012) 33:350–6.
doi: 10.2164/jandrol.111.014373
PubMed Abstract | Crossref Full Text | Google Scholar
140. Magras, IN, and Xenos, TD. RF radiation-induced changes in the prenatal development of mice. Bioelectromagnetics. (1997) 18:455–61. doi: 10.1002/(SICI)1521-186X(1997)18:6<455::AID-BEM8>3.0.CO;2-1 Crossref Full Text | Google Scholar
141. Panagopoulos, DJ, Karabarbounis, A, and Margaritis, LH. Effect of GSM 900-MHz Mobile phone radiation on the reproductive capacity of Drosophila melanogaster. Electromagn Biol Med. (2004) 23:29–43. doi: 10.1081/JBC-120039350 Crossref Full Text | Google Scholar
142. Panagopoulos, DJ, Chavdoula, ED, Karabarbounis, A, and Margaritis, LH. Comparison of bioactivity between GSM 900 MHz and DCS 1800 MHz Mobile telephony radiation. Electromagn Biol Med. (2007) 26:33–44. doi: 10.1080/15368370701205644
PubMed Abstract | Crossref Full Text | Google Scholar
143. Sharma, VP, and Kumar, NR. Changes in honeybee behaviour and biology under the influence of cellphone radiations. Curr Sci. (2010) 98:1376–8. Google Scholar
144. Wdowiak, A, Wdowiak, L, and Wiktor, H. Evaluation of the effect of using mobile phones on male fertility. Ann Agric Environ Med. (2007) 14:169–72. PubMed Abstract | Google Scholar
145. Irani, M, Aradmehr, M, Ghorbani, M, and Baghani, R. Electromagnetic field exposure and abortion in pregnant women: a systematic review and meta-analysis. Malays J Med Sci. (2023) 30:70–80.
doi: 10.21315/mjms2023.30.5.6
PubMed Abstract | Crossref Full Text | Google Scholar
146. Bacandritsos, N, Granato, A, Budge, G, Papanastasiou, I, Roinioti, E, Caldon, M, et al. Sudden deaths and colony population decline in Greek honey bee colonies. J Invertebr Pathol. (2010) 105:335–40.
doi: 10.1016/j.jip.2010.08.004
PubMed Abstract | Crossref Full Text | Google Scholar
147. Balmori, A. Possible effects of electromagnetic fields from phone masts on a population of White stork (Ciconia ciconia). Electromagn Biol Med. (2005) 24:109–19. doi: 10.1080/15368370500205472
Crossref Full Text | Google Scholar
148. Balmori, A, and Hallberg, O. The urban decline of the house sparrow (Passer domesticus): a possible link with electromagnetic radiation. Electromagn Biol Med. (2007) 26:141–51.
doi: 10.1080/15368370701410558
PubMed Abstract | Crossref Full Text | Google Scholar
149. Cucurachi, S, Tamis, WLM, Vijver, MG, Peijnenburg, WJGM, Bolte, JFB, and de Snoo, GR. A review of the ecological effects of radiofrequency electromagnetic fields (RF-EMF). Environ Int. (2013) 51:116–40. doi: 10.1016/j.envint.2012.10.009
PubMed Abstract | Crossref Full Text | Google Scholar
150. Everaert, J, and Bauwens, D. A possible effect of electromagnetic radiation from mobile phone base stations on the number of breeding house sparrows (Passer domesticus). Electromagn Biol Med. (2007) 26:63–72. doi: 10.1080/15368370701205693
PubMed Abstract | Crossref Full Text | Google Scholar
151. Batellier, F, Couty, I, Picard, D, and Brillard, JP. Effects of exposing chicken eggs to a cell phone in “call” position over the entire incubation period. Theriogenology. (2008) 69:737–45.
doi: 10.1016/j.theriogenology.2007.12.006
PubMed Abstract | Crossref Full Text | Google Scholar
152. Balmori, A. The incidence of electromagnetic pollution on the amphibian decline: is this an important piece of the puzzle? Toxicol Environ Chem. (2006) 88:287–99. doi: 10.1080/02772240600687200
Crossref Full Text | Google Scholar
153. Balmori, A. Mobile phone mast effects on common frog (Rana temporaria) tadpoles: the city turned into a laboratory. Electromagn Biol Med. (2010) 29:31–5. doi: 10.3109/15368371003685363
Crossref Full Text | Google Scholar
154. Ahlbom, A, Day, N, Feychting, M, Roman, E, Skinner, J, Dockerty, J, et al. A pooled analysis of magnetic fields and childhood leukaemia. Br J Cancer. (2000) 83:692–8. doi: 10.1054/bjoc.2000.1376
PubMed Abstract | Crossref Full Text | Google Scholar
155. Coghill, RW, Steward, J, and Philips, A. Extra low frequency electric and magnetic fields in the bed place of children diagnosed with leukaemia: a case-control study. Eur J Cancer Prev. (1996) 5:153–8.
doi: 10.1097/00008469-199606000-00002
PubMed Abstract | Crossref Full Text | Google Scholar
156. Coleman, MP, Bell, CM, Taylor, HL, and Primic-Zakelj, M. Leukaemia and residence near electricity transmission equipment: a case-control study. Br J Cancer. (1989) 60:793–8. doi: 10.1038/bjc.1989.362
PubMed Abstract | Crossref Full Text | Google Scholar
157. Draper, G, Vincent, T, Kroll, ME, and Swanson, J. Childhood cancer in relation to distance from high voltage power lines in England and Wales: a case-control study. BMJ. (2005) 330:1290.
doi: 10.1136/bmj.330.7503.1290
PubMed Abstract | Crossref Full Text | Google Scholar
158. Feychting, M, and Ahlbom, A. Magnetic fields and cancer in children residing near Swedish high – voltage power lines. Am J Epidemiol. (1993) 138:467–81. doi: 10.1093/oxfordjournals.aje.a116881
Crossref Full Text | Google Scholar
159. Feychting, M, and Ahlbom, A. Magnetic fields, leukemia and central nervous system tumors in Swedish adults residing near high – voltage power lines. Epidemiology. (1994) 5:501–9.
PubMed Abstract | Google Scholar
160. Greenland, S, Sheppard, AR, Kaune, WT, Poole, C, and Kelsh, MA. A pooled analysis of magnetic fields, wire codes, and childhood leukemia. Childhood leukemia-EMF study group. Epidemiology. (2000) 11:624–34. doi: 10.1097/00001648-200011000-00003
PubMed Abstract | Crossref Full Text | Google Scholar
161. Kheifets, L, Ahlbom, A, Crespi, CM, Draper, G, Hagihara, J, Lowenthal, RM, et al. Pooled analysis of recent studies on magnetic fields and childhood leukaemia. Br J Cancer. (2010) 103:1128–35.
Google Scholar
162. Miller, A, To, T, Agnew, DA, et al. Leukemia following occupational exposure to 60-Hz electric and magnetic fields among Ontario electric utility workers. Am J Epidemiol. (1996) 144:150–60.
doi: 10.1093/oxfordjournals.aje.a008902
PubMed Abstract | Crossref Full Text | Google Scholar
163. Savitz, DA, Wachtel, H, Barnes, F, John, EM, and Tvrdik, JG. Case-control study of childhood cancer and exposure to 60Hz magnetic fields. Am J Epidemiol. (1988) 128:21–38.
doi: 10.1093/oxfordjournals.aje.a114943
PubMed Abstract | Crossref Full Text | Google Scholar
164. Villeneuve, PJ, Agnew, DA, Miller, AB, and Corey, PN. Non-Hodgkin’s lymphoma among electric utility workers in Ontario: the evaluation of alternate indices of exposure to 60 Hz electric and magnetic fields. Occup Environ Med. (2000) 57:249–57. doi: 10.1136/oem.57.4.249
PubMed Abstract | Crossref Full Text | Google Scholar
165. Villeneuve, PJ, Agnew, DA, Miller, AB, Corey, PN, and Purdham, JT. Leukemia in electric utility workers: the evaluation of alternative indices of exposure to 60 Hz electric and magnetic fields. Am J Ind Med. (2000) 37:607–17. doi: 10.1002/(SICI)1097-0274(200006)37:6<607::AID-AJIM5>3.0.CO;2-L
PubMed Abstract | Crossref Full Text | Google Scholar
166. Wertheimer, N, and Leeper, E. Electrical wiring configurations and childhood cancer. Am J Epidemiol. (1979) 109:273–84. doi: 10.1093/oxfordjournals.aje.a112681
Crossref Full Text | Google Scholar
167. Wertheimer, N, and Leeper, E. Adult cancer related to electrical wires near the home. Int J Epidemiol. (1982) 11:345–55. doi: 10.1093/ije/11.4.345
PubMed Abstract | Crossref Full Text | Google Scholar
168. Hallberg, O, and Johansson, O. Melanoma incidence and frequency modulation (FM) broadcasting. Arch Environ Health. (2002) 57:32–40. doi: 10.1080/00039890209602914
PubMed Abstract | Crossref Full Text | Google Scholar
169. López, I, Félix, N, Rivera, M, Alonso, A, and Maestú, C. What is the radiation before 5G? A correlation study between measurements in situ and in real time and epidemiological indicators in Vallecas, Madrid. Environ Res. (2021) 194:110734. doi: 10.1016/j.envres.2021.110734
PubMed Abstract | Crossref Full Text | Google Scholar
170. Yakymenko, I, Sidorik, E, Kyrylenko, S, and Chekhun, V. Long-term exposure to microwave radiation provokes cancer growth: evidences from radars and mobile communication systems. Exp Oncol. (2011) 33:62–70.
PubMed Abstract | Google Scholar
171. Yakymenko, I, and Tsybulin, O. Carcinogenic effects of non-thermal exposure to wireless communication electromagnetic fields In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press (2022)
Google Scholar
172. Gulati, S, Yadav, A, Kumar, N, Kanupriya,, Aggarwal, NK, Kumar, R, et al. Effect of GSTM1 and GSTT1 polymorphisms on genetic damage in humans populations exposed to radiation from Mobile towers. Arch Environ Contam Toxicol. (2016) 70:615–25. doi: 10.1007/s00244-015-0195-y
PubMed Abstract | Crossref Full Text | Google Scholar
173. Zothansiama,, Zosangzuali, M, Lalramdinpuii, M, and Jagetia, GC. Impact of radiofrequency radiation on DNA damage and antioxidants in peripheral blood lymphocytes of humans residing in the vicinity of mobile phone base stations. Electromagn Biol Med. (2017) 36:295–305.
doi: 10.1080/15368378.2017.1350584
PubMed Abstract | Crossref Full Text | Google Scholar
174. Carlberg, M, and Hardell, L. Evaluation of Mobile phone and cordless phone use and glioma risk using the Bradford Hill viewpoints from 1965 on association or causation. Biomed Res Int. (2017) 2017:9218486–17. doi: 10.1155/2017/9218486
PubMed Abstract | Crossref Full Text | Google Scholar
175. Hardell, L, Carlberg, M, Söderqvist, F, Mild, KH, and Morgan, LL. Long-term use of cellular phones and brain tumours: increased risk associated with use for > or =10 years. Occup Environ Med. (2007) 64:626–32. doi: 10.1136/oem.2006.029751
PubMed Abstract | Crossref Full Text | Google Scholar
176. Hardell, L, and Carlberg, M. Mobile phones, cordless phones and the risk for brain tumours. Int J Oncol. (2009) 35:5–17. doi: 10.3892/ijo_00000307
PubMed Abstract | Crossref Full Text | Google Scholar
177. Hardell, L, Carlberg, M, and Hansson, MK. Epidemiological evidence for an association between use of wireless phones and tumor diseases. Pathophysiology. (2009) 16:113–22.
doi: 10.1016/j.pathophys.2009.01.003
PubMed Abstract | Crossref Full Text | Google Scholar
178. Hardell, L, Carlberg, M, Söderqvist, F, and Mild, KH. Case-control study of the association between malignant brain tumours diagnosed between 2007 and 2009 and mobile and cordless phone use. Int J Oncol. (2013) 43:1833–45. doi: 10.3892/ijo.2013.2111
PubMed Abstract | Crossref Full Text | Google Scholar
179. Hardell, L, Carlberg, M, and Hansson Mild, K. Use of mobile phones and cordless phones is associated with increased risk for glioma and acoustic neuroma. Pathophysiology. (2013) 20:85–110.
doi: 10.1016/j.pathophys.2012.11.001
PubMed Abstract | Crossref Full Text | Google Scholar
180. Hardell, L. Effects of Mobile phones on Children’s and Adolescents’ health: a commentary. Child Dev. (2018) 89:137–40. doi: 10.1111/cdev.12831
PubMed Abstract | Crossref Full Text | Google Scholar
181. Khurana, VG, Teo, C, Kundi, M, Hardell, L, and Carlberg, M. Cell phones and brain tumors: a review including the long-term epidemiologic data. Surg Neurol. (2009) 72:205–14.
doi: 10.1016/j.surneu.2009.01.019
PubMed Abstract | Crossref Full Text | Google Scholar
182. Miller, AB, Morgan, LL, Udasin, I, and Davis, DL. Cancer epidemiology update, following the 2011 IARC evaluation of radiofrequency electromagnetic fields (monograph 102). Environ Res. (2018) 167:673–83. doi: 10.1016/j.envres.2018.06.043
PubMed Abstract | Crossref Full Text | Google Scholar
183. Miller, AB, Sears, ME, Morgan, LL, Davis, DL, Hardell, L, Oremus, M, et al. Risks to health and well-being from radio-frequency radiation emitted by cell phones and other wireless devices. Front Public Health. (2019) 7:223. doi: 10.3389/fpubh.2019.00223
PubMed Abstract | Crossref Full Text | Google Scholar
184. Momoli, F, Siemiatycki, J, McBride, ML, Parent, MÉ, Richardson, L, Bedard, D, et al. Probabilistic multiple-bias modeling applied to the Canadian data from the interphone study of mobile phone use and risk of glioma, meningioma, acoustic neuroma, and parotid gland tumors. Am J Epidemiol. (2017) 186:885–93. doi: 10.1093/aje/kwx157
PubMed Abstract | Crossref Full Text | Google Scholar
185. Wang, Y, and Guo, X. Meta-analysis of association between mobile phone use and glioma risk. J Cancer Res Ther. (2016) 12:298–C300. doi: 10.4103/0973-1482.200759
PubMed Abstract | Crossref Full Text | Google Scholar
186. Navarro, A, et al. The microwave syndrome: a preliminary study in Spain. Electromagn Biol Med. (2003) 22:161–9. doi: 10.1081/JBC-120024625
Crossref Full Text | Google Scholar
187. Santini, R, Santini, P, Danze, JM, Le Ruz, P, and Seigne, M. Study of the health of people living in the vicinity of mobile phone base stations: I. Influences of distance and sex. Pathol Biol. (2002) 50:369–73.
doi: 10.1016/S0369-8114(02)00311-5
PubMed Abstract | Crossref Full Text | Google Scholar
188. Salama, OE, and Abou El Naga, RM. Cellular phones: are they detrimental? J Egypt Public Health Assoc. (2004) 79:197–223. Google Scholar
189. Abdel-Rassoul, G, El-Fateh, OA, Salem, MA, Michael, A, Farahat, F, El-Batanouny, M, et al. Neurobehavioral effects among inhabitants around mobile phone base stations. Neurotoxicology. (2007) 28:434–40. doi: 10.1016/j.neuro.2006.07.012
PubMed Abstract | Crossref Full Text | Google Scholar
190. Belpomme, D, and Irigaray, P. Electrohypersensitivity as a worldwide man-made electromagnetic pathology: a review of the medical evidence In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press (2022) Google Scholar
191. Blettner, M, Schlehofer, B, Breckenkamp, J, Kowall, B, Schmiedel, S, Reis, U, et al. Mobile phone base stations and adverse health effects: phase 1 of a population-based, cross-sectional study in Germany. Occup Environ Med. (2009) 66:118–23. doi: 10.1136/oem.2007.037721
PubMed Abstract | Crossref Full Text | Google Scholar
192. Davis, D, Birnbaum, L, Ben-Ishai, P, Taylor, H, Sears, M, Butler, T, et al. Wireless technologies, non-ionizing electromagnetic fields and children: identifying and reducing health risks. Curr Probl Pediatr Adolesc Health Care. (2023) 53:101374. doi: 10.1016/j.cppeds.2023.101374
PubMed Abstract | Crossref Full Text | Google Scholar
193. Greco, F, Garnier, O, Macioce, V, and Picot, MC. Prevalence of migraine disease in Electrohypersensitive patients. J Clin Med. (2023) 12:4092. doi: 10.3390/jcm12124092
PubMed Abstract | Crossref Full Text | Google Scholar
194. Hutter, H-P, Moshammer, H, Wallner, P, and Kundi, M. Subjective symptoms, sleeping problems, and cognitive performance in subjects living near mobile phone base stations. Occup Environ Med. (2006) 63:307–13. doi: 10.1136/oem.2005.020784
PubMed Abstract | Crossref Full Text | Google Scholar
195. Kundi, M, and Hutter, HP. Mobile phone base stations-effects on wellbeing and health. Pathophysiology. (2009) 16:123–35. doi: 10.1016/j.pathophys.2009.01.008
PubMed Abstract | Crossref Full Text | Google Scholar
196. Viel, JF, Clerc, S, Barrera, C, Rymzhanova, R, Moissonnier, M, Hours, M, et al. Residential exposure to radiofrequency fields from mobile phone base stations, and broadcast transmitters: a population-based survey with personal meter. Occup Environ Med. (2009) 66:550–6. doi: 10.1136/oem.2008.044180
PubMed Abstract | Crossref Full Text | Google Scholar
197. Irigaray, P, Caccamo, D, and Belpomme, D. Oxidative stress in electrohypersensitivity self-reporting patients: results of a prospective in vivo investigation with comprehensive molecular analysis. Int J Mol Med. (2018) 42:1885–98. doi: 10.3892/ijmm.2018.3774
PubMed Abstract | Crossref Full Text | Google Scholar
198. Hardell, L, and Nilsson, M. Summary of seven Swedish case reports on the microwave syndrome associated with 5G radiofrequency radiation. Rev Environ Health. (2024) 40:147–57.
doi: 10.1515/reveh-2024-0017
PubMed Abstract | Crossref Full Text | Google Scholar
199. Hardell, L, and Koppel, T. Spots with extremely high radiofrequency radiation after deployment of 5G base stations in Stockholm, Sweden. Ann Clin Med Case Rep. (2024) V14:1–8. Google Scholar
200. Lerchl, A, Klose, M, Grote, K, Wilhelm, AFX, Spathmann, O, Fiedler, T, et al. Tumor promotion by exposure to radiofrequency electromagnetic fields below exposure limits for humans. Biochem Biophys Res Commun. (2015) 459:585–90. doi: 10.1016/j.bbrc.2015.02.151
PubMed Abstract | Crossref Full Text | Google Scholar
201. Tillmann, T, Ernst, H, Streckert, J, Zhou, Y, Taugner, F, Hansen, V, et al. Indication of cocarcinogenic potential of chronic UMTS-modulated radiofrequency exposure in an ethylnitrosourea mouse model. Int J Radiat Biol. (2010) 86:529–41. doi: 10.3109/09553001003734501
PubMed Abstract | Crossref Full Text | Google Scholar
202. NTP. Toxicology and carcinogenesis studies in Hsd: Sprague Dawley SD rats exposed to whole-body radio frequency radiation at a frequency (900 MHz) and modulations (GSM and CDMA) used by cell phones. USA: NTP TR 595, Department of Health and Human Services (2018). Google Scholar
203. Falcioni, L, Bua, L, Tibaldi, E, Lauriola, M, de Angelis, L, Gnudi, F, et al. Report of final results regarding brain and heart tumors in Sprague-Dawley rats exposed from prenatal life until natural death to mobile phone radiofrequency field representative of a 1.8GHz GSM base station environmental emission. Environ Res. (2018) 165:496–503. doi: 10.1016/j.envres.2018.01.037
PubMed Abstract | Crossref Full Text | Google Scholar
204. IARC. Non-ionizing radiation, part 1: static and extremely low-frequency (ELF) electric and magnetic fields, vol. 80. Lyon, France: International Agency for Research on Cancer. (2002). Google Scholar
205. IARC. Non-ionizing radiation, part 2: radiofrequency electromagnetic fields, vol. 102. Lyon, France: International Agency for Research on Cancer. (2013). Google Scholar
206. Leach, V, Weller, S, and Redmayne, M. A novel database of bio-effects from non-ionizing radiation. Rev Environ Health. (2018) 33:273–80. doi: 10.1515/reveh-2018-0017
PubMed Abstract | Crossref Full Text | Google Scholar
207. Lerchl, A, Klose, M, and Drees, K. No increased DNA damage observed in the brain, liver, and lung of fetal mice treated with Ethylnitrosourea and exposed to UMTS radiofrequency electromagnetic fields. Bioelectromagnetics. (2020) 41:611–6. doi: 10.1002/bem.22301
PubMed Abstract | Crossref Full Text | Google Scholar
208. Manna, D, and Ghosh, R. Effect of radiofrequency radiation in cultured mammalian cells: a review. Electromagn Biol Med. (2016) 35:265–301. doi: 10.3109/15368378.2015.1092158
PubMed Abstract | Crossref Full Text | Google Scholar
209. Panagopoulos, DJ, Johansson, O, and Carlo, GL. Real versus simulated Mobile phone exposures in experimental studies. Biomed Res Int. (2015) 2015:607053. doi: 10.1155/2015/607053
PubMed Abstract | Crossref Full Text | Google Scholar
210. Panagopoulos, DJ, Yakymenko, I, and Chrousos, GP. Electromagnetic field-induced dysfunction of voltage-gated ion channels, oxidative stress, DNA damage, and related pathologies In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press (2022) Google Scholar
211. Schuermann, D, Ziemann, C, Barekati, Z, Capstick, M, Oertel, A, Focke, F, et al. Assessment of Genotoxicity in human cells exposed to modulated electromagnetic fields of wireless communication devices. Genes. (2020) 11:347. doi: 10.3390/genes11040347
PubMed Abstract | Crossref Full Text | Google Scholar
212. Verschaeve, L. Misleading scientific papers on health effects from wireless communication devices In: CD Geddes, editor. Microwave effects on DNA and proteins. Cham, Switzerland: Springer. (2017)
Google Scholar
213. Wood, A, Mate, R, and Karipidis, K. Meta-analysis of in vitro and in vivo studies of the biological effects of low-level millimetre waves. J Expo Sci Environ Epidemiol. (2021) 31:606–13.
doi: 10.1038/s41370-021-00307-7
PubMed Abstract | Crossref Full Text | Google Scholar
214. McCredden, JE, Cook, N, Weller, S, and Leach, V. Wireless technology is an environmental stressor requiring new understanding and approaches in health care. Front Public Health. (2022) 10:986315.
doi: 10.3389/fpubh.2022.986315
PubMed Abstract | Crossref Full Text | Google Scholar
215. McCredden, JE, Weller, S, and Leach, V. The assumption of safety is being used to justify the rollout of 5G technologies. Front Public Health. (2023) 11:1058454. doi: 10.3389/fpubh.2023.1058454
PubMed Abstract | Crossref Full Text | Google Scholar
216. Panagopoulos, DJ. Mobile telephony radiation effects on insect ovarian cells. The necessity for real exposures bioactivity assessment. The key role of polarization, and the ion forced-oscillation mechanism In: CD Geddes, editor. Microwave effects on DNA and proteins. Cham, Switzerland: Springer (2017) 1–48.
Google Scholar
217. Frank, JW. Electromagnetic fields, 5G and health: what about the precautionary principle? J Epidemiol Community Health. (2021) 75:562–6. doi: 10.1136/jech-2019-213595
PubMed Abstract | Crossref Full Text | Google Scholar
218. Hardell, L, and Nyberg, R. Appeals that matter or not on a moratorium on the deployment of the fifth generation, 5G, for microwave radiation. Mol Clin Oncol. (2020) 12:247–57. doi: 10.3892/mco.2020.1984
PubMed Abstract | Crossref Full Text | Google Scholar
219. Hardell, L, and Carlberg, M. Health risks from radiofrequency radiation, including 5G, should be assessed by experts with no conflicts of interest. Oncol Lett. (2020) 20:15. doi: 10.3892/ol.2020.11876
PubMed Abstract | Crossref Full Text | Google Scholar
220. Hardell, L, and Carlberg, M. Lost opportunities for cancer prevention: historical evidence on early warnings with emphasis on radiofrequency radiation. Rev Environ Health. (2021) 36:585–97.
doi: 10.1515/reveh-2020-0168
PubMed Abstract | Crossref Full Text | Google Scholar
221. Harremoes, P, Gee, D, MacGarvin, M, Stirling, A, Keys, J, Wynne, B, et al. The precautionary principle in the 20th century: late lessons from early warnings. London: Routledge (2013). Google Scholar
222. ICNIRP. Guidelines for limiting exposure to time-varying electric, magnetic, and electromagnetic fields (up to 300 GHz). Health Phys. (1998) 74:494–521. Google Scholar
223. ICNIRP. Guidelines for limiting exposure to time-varying electric and magnetic fields (1Hz to 100 kHz). Health Phys. (2010) 99:818–36. Google Scholar
224. ICNIRP. Guidelines for limiting exposure to electromagnetic fields (100 kHz to 300 GHz). Health Phys. (2020) 118:483–524. Google Scholar
225. Ishai, PB, Baldwin, HZ, Birnbaum, LS, Butler, T, Chamberlin, K, Davis, DL, et al. Applying the precautionary principle to wireless technology: policy dilemmas and systemic risks. Environ Sci Policy Sustain Dev. (2024) 66:5–18. doi: 10.1080/00139157.2024.2293631
Crossref Full Text | Google Scholar
226. Read, R, and O’Riordan, T. The precautionary principle under fire. Environ Sci Policy Sustain Dev. (2017) 59:4–15. doi: 10.1080/00139157.2017.1350005
Crossref Full Text | Google Scholar
227. Baan, R, Grosse, Y, Lauby-Secretan, B, El Ghissassi, F, Bouvard, V, et al. WHO International Agency for Research on Cancer monograph working group: carcinogenicity of radiofrequency electromagnetic fields. Lancet Oncol. (2011) 12:624–6. doi: 10.1016/S1470-2045(11)70147-4
Crossref Full Text | Google Scholar
228. Nyberg, NR, McCredden, JE, Weller, SG, and Hardell, L. The European Union prioritises economics over health in the rollout of radiofrequency technologies. Rev Environ Health. (2024) 39:47–64. doi: 10.1515/reveh-2022-0106
PubMed Abstract | Crossref Full Text | Google Scholar
229. Francis, G. Ionization phenomena in gases. London: Butterworths Scientific Publications (1960).
Google Scholar
230. Gomer, R. Field emission and field ionization. USA: Harvard University Press (1961).
Google Scholar
231. Alberts, B, Bray, D, Lewis, J, Raff, M, Roberts, K, and Watson, JD. Molecular biology of the cell. New York, USA: Garland Publishing, Inc (1994). Google Scholar
232. Finkel, T. Signal transduction by reactive oxygen species. J Cell Biol. (2011) 194:7–15.
doi: 10.1083/jcb.201102095
PubMed Abstract | Crossref Full Text | Google Scholar
233. Görlach, A, Bertram, K, Hudecova, S, and Krizanova, O. Calcium and ROS: a mutual interplay. Redox Biol. (2015) 6:260–71. doi: 10.1016/j.redox.2015.08.010
PubMed Abstract | Crossref Full Text | Google Scholar
234. Stryer, L. Biochemistry. New York: Freeman (1995). Google Scholar
235. Inoue, M, Sato, EF, Nishikawa, M, Park, AM, Kira, Y, Imada, I, et al. Mitochondrial generation of reactive oxygen species and its role in aerobic life. Curr Med Chem. (2003) 10:2495–505.
doi: 10.2174/0929867033456477
PubMed Abstract | Crossref Full Text | Google Scholar
236. Lushchak, VI. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem Biol Interact. (2014) 224:164–75. doi: 10.1016/j.cbi.2014.10.016
Crossref Full Text | Google Scholar
237. Ischiropoulos, H, Zhu, L, and Beckman, JS. Peroxynitrite formation from macrophage-derived nitric oxide. Arch Biochem Biophys. (1992) 298:446–51. doi: 10.1016/0003-9861(92)90433-W
Crossref Full Text | Google Scholar
238. Pall, ML. Electromagnetic fields act via activation of voltage-gated calcium channels to produce beneficial or adverse effects. J Cell Mol Med. (2013) 17:958–65. doi: 10.1111/jcmm.12088
PubMed Abstract | Crossref Full Text | Google Scholar
239. Schuermann, D, and Mevissen, M. Manmade electromagnetic fields and oxidative stress—biological effects and consequences for health. Int J Mol Sci. (2021) 22:3772. doi: 10.3390/ijms22073772
PubMed Abstract | Crossref Full Text | Google Scholar
240. Checa, J, and Aran, JM. Reactive oxygen species: drivers of physiological and pathological processes. J Inflamm Res. (2020) 13:1057–73. doi: 10.2147/JIR.S275595
PubMed Abstract | Crossref Full Text | Google Scholar
241. Andrés, CMC, Pérez de la Lastra, JM, Andrés Juan, C, Plou, FJ, and Pérez-Lebeña, E. Superoxide anion chemistry—its role at the core of the innate immunity. Int J Mol Sci. (2023) 24:1841.
doi: 10.3390/ijms24031841
PubMed Abstract | Crossref Full Text | Google Scholar
242. Koppers, AJ, De Iuliis, GN, Finnie, JM, McLaughlin, EA, and Aitken, RJ. Significance of mitochondrial reactive oxygen species in the generation of oxidative stress in spermatozoa. J Clin Endocrinol Metabol. (2008) 93:3199–207. doi: 10.1210/jc.2007-2616
PubMed Abstract | Crossref Full Text | Google Scholar
243. Gibb, Z, Griffin, RA, Aitken, RJ, and De Iuliis, GN. Functions and effects of reactive oxygen species in male fertility. Anim Reprod Sci. (2020) 220:106456. doi: 10.1016/j.anireprosci.2020.106456
PubMed Abstract | Crossref Full Text | Google Scholar
244. Brookes, PS, Yoon, Y, Robotham, JL, Anders, MW, and Sheu, SS. Calcium, ATP, and ROS: a mitochondrial love-hate triangle. Am J Physiol Cell Physiol. (2004) 287:C817–33.
doi: 10.1152/ajpcell.00139.2004 Crossref Full Text | Google Scholar
245. Bertero, E, and Maack, C. Calcium signaling and reactive oxygen species in mitochondria. Circ Res. (2018) 122:1460–78. doi: 10.1161/CIRCRESAHA.118.310082
Crossref Full Text | Google Scholar
246. Lowe, SW, and Lin, AW. Apoptosis in cancer. Carcinogenesis. (2000) 21:485–95.
doi: 10.1093/carcin/21.3.485
PubMed Abstract | Crossref Full Text | Google Scholar
247. Bedard, K, and Krause, KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev. (2007) 87:245–313. doi: 10.1152/physrev.00044.2005
PubMed Abstract | Crossref Full Text | Google Scholar
248. Gamaley, Ι, Augsten, Κ, and Berg, Η. Electrostimulation of macrophage NADPH oxidase by modulated high-frequency electromagnetic fields. Bioelectrochem Bioenerg. (1995) 38:415–8.
doi: 10.1016/0302-4598(95)01836-4 Crossref Full Text | Google Scholar
249. Iverson, D, De Chatelet, LR, Spitznagel, JK, and Wang, P. Comparison of NADH and NADPH oxidase activities in granules isolated from human polymorphonuclear leukocytes with a fluorometric assay. J Clin Invest. (1977) 59:282–90. doi: 10.1172/JCI108639
PubMed Abstract | Crossref Full Text | Google Scholar
250. Li, JM, and Shah, AM. Differential NADPH- versus NADH-dependent superoxide production by phagocyte-type endothelial cell NADPH oxidase. Cardiovasc Res. (2001) 52:477–86.
doi: 10.1016/s0008-6363(01)00407-2
PubMed Abstract | Crossref Full Text | Google Scholar
251. Panday, A, Sahoo, MK, Osorio, D, and Batra, S. NADPH oxidases: an overview from structure to innate immunity-associated pathologies. Cell Mol Immunol. (2015) 12:5–23. doi: 10.1038/cmi.2014.89
PubMed Abstract | Crossref Full Text | Google Scholar
252. Cross, AR, and Segal, AW. The NADPH oxidase of professional phagocytes—prototype of the NOX electron transport chain systems. Biochim Biophys Acta. (2004) 1657:1–22.
doi: 10.1016/j.bbabio.2004.03.008
PubMed Abstract | Crossref Full Text | Google Scholar
253. Henderson, LM. NADPH oxidase subunit gp91phox: a proton pathway. Protoplasma. (2001) 217:37–42. doi: 10.1007/BF01289411 PubMed Abstract | Crossref Full Text | Google Scholar
254. Musset, B, Cherny, VV, Morgan, D, and De Coursey, TE. The intimate and mysterious relationship between proton channels and NADPH oxidase. FEBS Lett. (2009) 583:7–12.
doi: 10.1016/j.febslet.2008.12.005
PubMed Abstract | Crossref Full Text | Google Scholar
255. André-Lévigne, D, Modarressi, A, Pepper, MS, et al. Reactive oxygen species and NOX enzymes are emerging as key players in cutaneous wound repair. Int J Mol Sci. (2017) 18:2149.
doi: 10.3390/ijms18102149
PubMed Abstract | Crossref Full Text | Google Scholar
256. Fang, J, Sheng, R, and Qin, Z-H. NADPH oxidases in the central nervous system: regional and cellular localization and the possible link to brain diseases. Antioxid Redox Signal. (2021) 35:951–73.
doi: 10.1089/ars.2021.0040
PubMed Abstract | Crossref Full Text | Google Scholar
257. Friedman, J, Kraus, S, Hauptman, Y, Schiff, Y, and Seger, R. Mechanism of short-term ERK activation by electromagnetic fields at mobile phone frequencies. Biochem J. (2007) 405:559–68.
doi: 10.1042/BJ20061653
PubMed Abstract | Crossref Full Text | Google Scholar
258. Pacher, P, Beckman, JS, and Liaudet, L. Nitric oxide and Peroxynitrite in health and disease. Physiol Rev. (2007) 87:315–424. doi: 10.1152/physrev.00029.2006
PubMed Abstract | Crossref Full Text | Google Scholar
259. Pilla, AA. Electromagnetic fields instantaneously modulate nitric oxide signaling in challenged biological systems. Biochem Biophys Res Commun. (2012) 426:330–3. doi: 10.1016/j.bbrc.2012.08.078
PubMed Abstract | Crossref Full Text | Google Scholar
260. Lai, H, and Singh, NP. Magnetic-field-induced DNA strand breaks in brain cells of the rat. Environ Health Perspect. (2004) 112:687–94. doi: 10.1289/ehp.6355
Crossref Full Text | Google Scholar
261. Moon, HK, Yang, ES, and Park, JW. Protection of peroxynitrite-induced DNA damage by dietary antioxidants. Arch Pharm Res. (2006) 29:213–7. doi: 10.1007/BF02969396
PubMed Abstract | Crossref Full Text | Google Scholar
262. Sakihama, Y, Maeda, M, Hashimoto, M, Tahara, S, and Hashidoko, Y. Beetroot betalain inhibits peroxynitrite- mediated tyrosine nitration and DNA strand damage. Free Radic Res. (2012) 46:93–9.
doi: 10.3109/10715762.2011.641157
PubMed Abstract | Crossref Full Text | Google Scholar
263. Förstermann, U, and Sessa, WC. Nitric oxide synthases: regulation and function. Eur Heart J. (2012) 33:829–37. doi: 10.1093/eurheartj/ehr304
PubMed Abstract | Crossref Full Text | Google Scholar
264. Bortolotti, M, Polito, L, Battelli, MG, and Bolognesi, A. Xanthine oxidoreductase: one enzyme for multiple physiological tasks. Redox Biol. (2021) 41:101882. doi: 10.1016/j.redox.2021.101882
PubMed Abstract | Crossref Full Text | Google Scholar
265. Manikandan, P, and Nagini, S. Cytochrome P450 structure, function and clinical significance: a review. Curr Drug Targets. (2018) 19:38–54. doi: 10.2174/1389450118666170125144557
PubMed Abstract | Crossref Full Text | Google Scholar
266. Becchetti, A. Ion channels and transporters in cancer. 1. Ion channels and cell proliferation in cancer. Am J Phys Cell Phys. (2011) 301:C255–65. Google Scholar
267. Lang, F, Föller, M, Lang, KS, Lang, PA, Ritter, M, Gulbins, E, et al. Ion channels in cell proliferation and apoptotic cell death. J Membr Biol. (2005) 205:147–57. doi: 10.1007/s00232-005-0780-5
Crossref Full Text | Google Scholar
268. Chattipakorn, N, Kumfu, S, Fucharoen, S, and Chattipakorn, S. Calcium channels and iron uptake into the heart. World J Cardiol. (2011) 3:215–8. doi: 10.4330/wjc.v3.i7.215
PubMed Abstract | Crossref Full Text | Google Scholar
269. Gaasch, JA, Geldenhuys, WJ, Lockman, PR, Allen, DD, and Van der Schyf, CJ. Voltage-gated calcium channels provide an alternate route for Iron uptake in neuronal cell cultures. Neurochem Res. (2007) 32:1686–93. doi: 10.1007/s11064-007-9313-1
PubMed Abstract | Crossref Full Text | Google Scholar
270. Salsbury, G, Cambridge, EL, McIntyre, Z, Arends, MJ, Karp, NA, Isherwood, C, et al. Disruption of the potassium channel regulatory subunit KCNE2 causes iron-deficient anemia. Exp Hematol. (2014) 42:1053–1058.e1. doi: 10.1016/j.exphem.2014.07.269
PubMed Abstract | Crossref Full Text | Google Scholar
271. Alberts, B, Johnson, A, Lewis, J, Raff, M, Roberts, K, and Walter, P. Membrane transport of small molecules and the electrical properties of membranes-molecular biology of the cell. New York, USA: Garland Science (2002). Google Scholar
272. Blank, M, and Soo, L. Ion activation of the Na, K-ATPase in alternating currents. Bioelectrochem Bioenerg. (1990) 24:51–61. doi: 10.1016/0302-4598(80)85006-9
Crossref Full Text | Google Scholar
273. Serpersu, EH, and Tsong, TY. Activation of electrogenic Rb transport of (Na/K)-ATPase by an electric field. J Biol Chem. (1984) 259:7155–62. doi: 10.1016/S0021-9258(17)39851-4
Crossref Full Text | Google Scholar
274. Pratt, RD, Brickman, CR, Cottrill, CL, Shapiro, JI, and Liu, J. The Na/K-ATPase signaling: from specific ligands to general reactive oxygen species. Int J Mol Sci. (2018) 19:2600. doi: 10.3390/ijms19092600
PubMed Abstract | Crossref Full Text | Google Scholar
275. Zhang, L, Zhang, Z, Guo, H, and Wang, Y. Na+/K+-ATPase-mediated signal transduction and Na+/K+-ATPase regulation. Fundam Clin Pharmacol. (2008) 22:615–21. doi: 10.1111/j.1472-8206.2008.00620.x
PubMed Abstract | Crossref Full Text | Google Scholar
276. Prasad, KN. Handbook of radiobiology. 2nd ed. Boca Raton: CRC Press (1995). Google Scholar
277. Barzilai, A, and Yamamoto, K. DNA damage responses to oxidative stress. DNA Repair. (2004) 3:1109–15. doi: 10.1016/j.dnarep.2004.03.002
PubMed Abstract | Crossref Full Text | Google Scholar
278. Cooke, MS, Evans, MD, Dizdaroglu, M, and Lunec, J. Oxidative DNA damage: mechanisms, mutation, and disease. FASEB J. (2003) 17:1195–214. doi: 10.1096/fj.02-0752rev
PubMed Abstract | Crossref Full Text | Google Scholar
279. Sillar, JR, Germon, ZP, De Iuliis, GN, and Dun, MD. The role of reactive oxygen species in acute myeloid leukaemia. Int J Mol Sci. (2019) 20:6003. doi: 10.3390/ijms20236003
PubMed Abstract | Crossref Full Text | Google Scholar
280. Valko, M, Leibfritz, D, Moncol, J, Cronin, MTD, Mazur, M, and Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. (2007) 39:44–84.
doi: 10.1016/j.biocel.2006.07.001
PubMed Abstract | Crossref Full Text | Google Scholar
281. Burney, S, Caulfield, JL, Niles, JC, Wishnok, JS, and Tannenbaum, SR. The chemistry of DNA damage from nitric oxide and peroxynitrite. Mutat Res. (1999) 424:37–49. doi: 10.1016/S0027-5107(99)00006-8
PubMed Abstract | Crossref Full Text | Google Scholar
282. Yu, H, Venkatarangan, L, Wishnok, JS, and Tannenbaum, SR. Quantitation of four guanine oxidation products from reaction of DNA with varying doses of Peroxynitrite. Chem Res Toxicol. (2005) 18:1849–57. doi: 10.1021/tx050146h PubMed Abstract | Crossref Full Text | Google Scholar
283. Szabo, G, and Baehrle, S. Role of nitrosative stress and poly(ADP-ribose) polymerase activation in myocardial reperfusion injury. Curr Vasc Pharmacol. (2005) 3:215–20. doi: 10.2174/1570161054368599
PubMed Abstract | Crossref Full Text | Google Scholar
284. Lymar, SV, Khairutdinov, RF, and Hurst, JK. Hydroxyl radical formation by O-O bond homolysis in peroxynitrous acid. Inorg Chem. (2003) 42:5259–66. doi: 10.1021/ic030104l
PubMed Abstract | Crossref Full Text | Google Scholar
285. Pérez de la Lastra, JM, Juan, CA, Plou, FJ, and Pérez-Lebeña, E. The nitration of proteins, lipids and DNA by peroxynitrite derivatives-chemistry involved and biological relevance. Stress. (2022) 2:53–64.
doi: 10.3390/stresses2010005 Crossref Full Text | Google Scholar
286. De Coursey, T, Morgan, D, and Cherny, V. The voltage dependence of NADPH oxidase reveals why phagocytes need proton channels. Nature. (2003) 422:531–4. doi: 10.1038/nature01523
PubMed Abstract | Crossref Full Text | Google Scholar
287. Balasubramanian, B, Pogozelski, WK, and Tullius, TD. DNA strand breaking by hydroxyl radical is governed by the accessible surface area of the hydrogen atom of the DNA backbone. PNAS. (1998) 95:9738–43. doi: 10.1073/pnas.95.17.9738 Crossref Full Text | Google Scholar
288. Cadet, J, Delatour, T, Douki, T, Gasparutto, D, Pouget, JP, Ravanat, JL, et al. Hydroxyl radicals and DNA base damage. Mutat Res. (1999) 424:9–21. doi: 10.1016/S0027-5107(99)00004-4
PubMed Abstract | Crossref Full Text | Google Scholar
289. Cadet, J, and Wagner, JR. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb Perspect Biol. (2013) 5:a012559. doi: 10.1101/cshperspect.a012559
PubMed Abstract | Crossref Full Text | Google Scholar
290. Halliwell, B. Biochemistry of oxidative stress. Biochem Soc Trans. (2007) 35:1147–50.
doi: 10.1042/BST0351147 Crossref Full Text | Google Scholar
291. Tsunoda, M, Sakaue, T, Naito, S, Sunami, T, Abe, N, Ueno, Y, et al. Insights into the structures of DNA damaged by hydroxyl radical: crystal structures of DNA duplexes containing 5-formyluracil. J Nucleic Acids. (2010) 2010:107289. doi: 10.4061/2010/107289
PubMed Abstract | Crossref Full Text | Google Scholar
292. Fenton, HJH. Oxidation of tartaric acid in presence of iron. J Chem Soc Trans. (1894) 65:899–910.
doi: 10.1039/CT8946500899 Crossref Full Text | Google Scholar
293. Valko, M, Rhodes, CJ, Moncol, J, Izakovic, M, and Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. (2006) 160:1–40. doi: 10.1016/j.cbi.2005.12.009
PubMed Abstract | Crossref Full Text | Google Scholar
294. Casarett, AP. Radiation biology. New Jersey: Prentice-Hall Inc (1968). Google Scholar
295. Coggle, JE. Biological effects of radiation. London: Taylor & Francis (1983). Google Scholar
296. Abolfath, RM, Van Duin, ACT, and Brabec, T. Reactive molecular dynamics study on the first steps of DNA damage by free hydroxyl radicals. Chem A Eur J. (2011) 115:11045–9. doi: 10.1021/jp204894m
PubMed Abstract | Crossref Full Text | Google Scholar
297. Panagopoulos, DJ, Karabarbounis, A, and Chrousos, GP. Biophysical mechanism of animal magnetoreception, orientation and navigation. Sci Rep. (2024) 14:30053.
doi: 10.1038/s41598-024-77883-9 PubMed Abstract | Crossref Full Text | Google Scholar
298. Bertagna, F, Lewis, R, Silva, SRP, McFadden, J, and Jeevaratnam, K. Effects of electromagnetic fields on neuronal ion channels: a systematic review. Ann N Y Acad Sci. (2021) 1499:82–103.
doi: 10.1111/nyas.14597 PubMed Abstract | Crossref Full Text | Google Scholar
299. Liman, ER, Hess, P, Weaver, F, and Koren, G. Voltage sensing residues in the S4 region of a mammalian K+ channel. Nature. (1991) 353:752–6. doi: 10.1038/353752a0
PubMed Abstract | Crossref Full Text | Google Scholar
300. Miller, C. An overview of the potassium channel family. Genome Biol. (2000) 1:reviews0004.1.
doi: 10.1186/gb-2000-1-4-reviews0004
PubMed Abstract | Crossref Full Text | Google Scholar
301. Noda, M, Ikeda, T, Kayano, T, Suzuki, H, Takeshima, H, Kurasaki, M, et al. Existence of distinct sodium channel messenger RNAs in rat brain. Nature. (1986) 320:188–92. doi: 10.1038/320188a0
PubMed Abstract | Crossref Full Text | Google Scholar
302. Panagopoulos, DJ. Mechanism of ion forced-oscillation and voltage-gated ion channel dysfunction by polarized and coherent electromagnetic fields In: DJ Panagopoulos, editor. Electromagnetic fields of wireless communications: biological and health effects. Boca Raton: CRC Press, Taylor and Francis (2022)
Google Scholar
303. Sandipan, C, and Baron, C. Basic mechanisms of voltage sensing In: J Zheng and MC Trudeau, editors. Handbook of ion channels. Boca Raton: CRC Press (2015) Google Scholar
304. Zhang, XC, Yang, H, Liu, Z, and Sun, F. Thermodynamics of voltage-gated ion channels. Biophys Rep. (2018) 4:300–19. doi: 10.1007/s41048-018-0074-y
PubMed Abstract | Crossref Full Text | Google Scholar
305. Panagopoulos, DJ, Messini, N, Karabarbounis, A, Filippetis, AL, and Margaritis, LH. A mechanism for action of oscillating electric fields on cells. Biochem Biophys Res Commun. (2000) 272:634–40. doi: 10.1006/bbrc.2000.2746 PubMed Abstract | Crossref Full Text | Google Scholar
306. Panagopoulos, DJ, Karabarbounis, A, and Margaritis, LH. Mechanism for action of electromagnetic fields on cells. Biochem Biophys Res Commun. (2002) 298:95–102. doi: 10.1016/S0006-291X(02)02393-8
Crossref Full Text | Google Scholar
307. Balcavage, WX, Alvager, T, Swez, J, Goff, CW, Fox, MT, Abdullyava, S, et al. A mechanism for action of extremely low frequency electromagnetic fields on biological systems. Biochem Biophys Res Commun. (1996) 222:374–8. doi: 10.1006/bbrc.1996.0751
PubMed Abstract | Crossref Full Text | Google Scholar
308. Honig, BH, Hubbell, WL, and Flewelling, RF. Electrostatic interactions in membranes and proteins. Ann Rev Biophys Biophys Chem. (1986) 15:163–93. doi: 10.1146/annurev.bb.15.060186.001115
PubMed Abstract | Crossref Full Text | Google Scholar
309. Cleary, SF, Liu, LM, Graham, R, and Diegelmann, RF. Modulation of tendon fibroplasia by exogenous electric currents. Bioelectromagnetics. (1988) 9:183–94. doi: 10.1002/bem.2250090210
PubMed Abstract | Crossref Full Text | Google Scholar
310. McLeod, KJ, Lee, RC, and Ehrlich, HP. Frequency dependence of electric field modulation of fibroblast protein synthesis. Science. (1987) 236:1465–9. doi: 10.1126/science.3589667
PubMed Abstract | Crossref Full Text | Google Scholar
311. England, SJ, and Robert, D. The ecology of electricity and electroreception. Biol Rev. (2022) 97:383–413. doi: 10.1111/brv.12804 PubMed Abstract | Crossref Full Text | Google Scholar
312. Johnsen, S, and Lohmann, KJ. Magnetoreception in animals. Phys Today. (2008) 61:29–35.
doi: 10.1063/1.2897947 Crossref Full Text | Google Scholar
313. Cecchetto, C, Maschietto, M, Boccaccio, P, and Vassanelli, S. Electromagnetic field affects the voltage-dependent potassium channel Kv1.3. Electromagn Biol Med. (2020) 39:316–22.
doi: 10.1080/15368378.2020.1799386 PubMed Abstract | Crossref Full Text | Google Scholar
314. El-Swefy, S, Soliman, H, and Huessein, M. Calcium channel blockade alleviates brain injury induced by long-term exposure to an electromagnetic field. J Appl Biomed. (2008) 6:153–63.
doi: 10.32725/jab.2008.019 Crossref Full Text | Google Scholar
315. Liburdy, RP. Calcium signaling in lymphocytes and ELF fields. FEBS Lett. (1992) 301:53–9.
doi: 10.1016/0014-5793(92)80209-Y
PubMed Abstract | Crossref Full Text | Google Scholar
316. Piacentini, R, Ripoli, C, Mezzogori, D, Azzena, GB, and Grassi, C. Extremely low-frequency electromagnetic fields promote in vitro neurogenesis via upregulation of Cav1-channel activity. J Cell Physiol. (2008) 215:129–39. doi: 10.1002/jcp.21293
PubMed Abstract | Crossref Full Text | Google Scholar
317. Zheng, Y, Xia, P, Dong, L, Tian, L, and Xiong, C. Effects of modulation on sodium and potassium channel currents by extremely low frequency electromagnetic fields stimulation on hippocampal CA1 pyramidal cells. Electromagn Biol Med. (2021) 40:274–85. doi: 10.1080/15368378.2021.1885433
PubMed Abstract | Crossref Full Text | Google Scholar
318. Orfali, R, Alwatban, AZ, Orfali, RS, Lau, L, Chea, N, Alotaibi, AM, et al. Oxidative stress and ion channels in neurodegenerative diseases. Front Physiol. (2024) 15:1320086.
doi: 10.3389/fphys.2024.1320086 PubMed Abstract | Crossref Full Text | Google Scholar
319. Batcioglu, K, Uyumlu, AB, Satilmis, B, Yildirim, B, Yucel, N, Demirtas, H, et al. Oxidative stress in the in vivo DMBA rat model of breast cancer: suppression by a voltage-gated Sodium Channel inhibitor (RS100642). Basic Clin Pharmacol Toxicol. (2012) 111:137–41. doi: 10.1111/j.1742-7843.2012.00880.x
PubMed Abstract | Crossref Full Text | Google Scholar
320. Afanas’ev, I. New nucleophilic mechanisms of ROS-dependent epigenetic modifications: comparison of aging and cancer. Aging Dis. (2014) 5:52–62. doi: 10.14336/AD.2014.050052
PubMed Abstract | Crossref Full Text | Google Scholar
321. Ramírez, A, Vázquez-Sánchez, AY, Carrión-Robalino, N, and Camacho, J. Ion channels and oxidative stress as a potential link for the diagnosis or treatment of liver diseases. Oxidative Med Cell Longev. (2016) 2016:3928714. doi: 10.1155/2016/3928714 PubMed Abstract | Crossref Full Text | Google Scholar
322. O’Hare Doig, RL, Chiha, W, Giacci, MK, Yates, NJ, Bartlett, CA, Smith, NM, et al. Specific ion channels contribute to key elements of pathology during secondary degeneration following neurotrauma. BMC Neurosci. (2017) 18:62. doi: 10.1186/s12868-017-0380-1
PubMed Abstract | Crossref Full Text | Google Scholar
323. Esteras, N, Kundel, F, Amodeo, GF, Pavlov, EV, Klenerman, D, and Abramov, AY. Insoluble tau aggregates induce neuronal death through modification of membrane ion conductance, activation of voltage-gated calcium channels and NADPH oxidase. FEBS J. (2021) 288:127–41. doi: 10.1111/febs.15340
PubMed Abstract | Crossref Full Text | Google Scholar
324. Fouda, MA, Ghovanloo, MR, and Ruben, PC. Cannabidiol protects against high glucose-induced oxidative stress and cytotoxicity in cardiac voltage-gated sodium channels. Br J Pharmacol. (2020) 177:2932–46. doi: 10.1111/bph.15020 PubMed Abstract | Crossref Full Text | Google Scholar
325. Thomas, MP, Chartrand, K, Reynolds, A, Vitvitsky, V, Banerjee, R, and Gendelman, HE. Ion channel blockade attenuates aggregated alpha synuclein induction of microglial reactive oxygen species: relevance for the pathogenesis of Parkinson’s disease. J Neurochem. (2007) 100:503–19.
doi: 10.1111/j.1471-4159.2006.04315.x
PubMed Abstract | Crossref Full Text | Google Scholar
326. Li, M, and Lester, HA. Ion Channel diseases of the central nervous system. CNS Drug Rev. (2001) 7:214–40. doi: 10.1111/j.1527-3458.2001.tb00196.x
PubMed Abstract | Crossref Full Text | Google Scholar
327. Huang, H, and Shakkottai, VG. Targeting ion channels and Purkinje neuron intrinsic membrane excitability as a therapeutic strategy for cerebellar Ataxia. Life. (2023) 13:1350. doi: 10.3390/life13061350
PubMed Abstract | Crossref Full Text | Google Scholar
328. Kourie, JI. Interaction of reactive oxygen species with ion transport mechanisms. Am J Phys. (1998) 275:C1–C24. Google Scholar
329. Lombardi, AA, Gibb, AA, Arif, E, Kolmetzky, DW, Tomar, D, Luongo, TS, et al. Mitochondrial calcium exchange links metabolism with the epigenome to control cellular differentiation. Nat Commun. (2019) 10:1–17. doi: 10.1038/s41467-019-12103-x PubMed Abstract | Crossref Full Text | Google Scholar
330. Patergnani, S, Danese, A, Bouhamida, E, Aguiari, G, Previati, M, Pinton, P, et al. Various aspects of calcium signaling in the regulation of apoptosis, autophagy, cell proliferation, and cancer. Int J Mol Sci. (2020) 21:8323. doi: 10.3390/ijms21218323 PubMed Abstract | Crossref Full Text | Google Scholar
331. Akbarali, HI. Oxidative stress and ion channels In: I Lahers, editor. Systems biology of free radicals and antioxidants. Berlin, Heidelberg: Springer. (2014) 355–73. Google Scholar
332. Nitahara, JA, Cheng, W, Liu, Y, Li, B, Leri, A, Li, P, et al. Intracellular calcium, DNase activity and myocyte apoptosis in aging Fischer 344 rats. J Mol Cell Cardiol. (1998) 30:519–35.
doi: 10.1006/jmcc.1997.0616 PubMed Abstract | Crossref Full Text | Google Scholar
333. Bawin, SM, and Adey, WR. Sensitivity of calcium binding in cerebral tissue to weak environmental electric fields oscillating at low frequency. Proc Natl Acad Sci USA. (1976) 73:1999–2003.
Google Scholar
334. Barbier, E, Vetret, B, and Dufy, B. Stimulation of Ca2+ influx in rat pituitary cells under exposure to a 50 Hz magnetic field. Bioelectromagnetics. (1996) 17:303–11.
doi: 10.1002/(SICI)1521-186X(1996)17:4<303::AID-BEM6>3.0.CO;2-7
PubMed Abstract | Crossref Full Text | Google Scholar
335. Dutta, SK, Subramaniam, A, Ghosh, B, and Parshad, R. Microwave radiation – induced calcium ion efflux from human neuroblastoma cells in culture. Bioelectromagnetics. (1984) 5:71–8.
doi: 10.1002/bem.2250050108 PubMed Abstract | Crossref Full Text | Google Scholar
336. Gobba, F, Malagoli, D, and Ottaviani, E. Effects of 50 Hz magnetic fields on fMLP-induced shape changes in invertebrate immunocytes: the role of calcium ion channels. Bioelectromagnetics. (2003) 24:277–82. doi: 10.1002/bem.10102 PubMed Abstract | Crossref Full Text | Google Scholar
337. Grassi, C, D’Ascenzo, M, Torsello, A, Martinotti, G, Wolf, F, Cittadini, A, et al. Effects of 50 Hz electromagnetic fields on voltage-gated Ca2+ channels and their role in modulation of neuroendocrine cell proliferation and death. Cell Calcium. (2004) 35:307–15. doi: 10.1016/j.ceca.2003.09.001
Crossref Full Text | Google Scholar
338. Jeong, JH, Kum, C, Choi, HJ, Park, ES, and Sohn, UD. Extremely low frequency magnetic field induces hyperalgesia in mice modulated by nitric oxide synthesis. Life Sci. (2006) 78:1407–12.
doi: 10.1016/j.lfs.2005.07.006 PubMed Abstract | Crossref Full Text | Google Scholar
339. Lisi, A, Ledda, M, Rosola, E, Pozzi, D, Emilia, ED′, Giuliani, L, et al. Extremely low frequency electromagnetic field exposure promotes differentiation of pituitary corticotrope-derived AtT20 D16V cells. Bioelectromagnetics. (2006) 27:641–51. doi: 10.1002/bem.20255
PubMed Abstract | Crossref Full Text | Google Scholar
340. Marchionni, I, Paffi, A, Pellegrino, M, Liberti, M, Apollonio, F, Abeti, R, et al. Comparison between low-level 50 Hz and 900 MHz electromagnetic stimulation on single channel ionic currents and on firing frequency in dorsal root ganglion isolated neurons. Biochim Biophys Acta. (2006) 1758:597–605.
doi: 10.1016/j.bbamem.2006.03.014 PubMed Abstract | Crossref Full Text | Google Scholar
341. Morgado-Valle, C, Verdugo-Dıaz, L, Garcıa, DE, et al. The role of voltage-gated Ca2+ channels in neurite growth of cultured chromaffin cells induced by extremely low frequency (ELF) magnetic field stimulation. Cell Tissue Res. (1998) 291:217–30. doi: 10.1007/s004410050992
PubMed Abstract | Crossref Full Text | Google Scholar
342. De Coursey, TE. Interactions between NADPH oxidase and voltage-gated proton channels: why electron transport depends on proton transport. FEBS Lett. (2003) 555:57–61.
doi: 10.1016/S0014-5793(03)01103-7 PubMed Abstract | Crossref Full Text | Google Scholar
343. Seredenina, T, Demaurex, N, and Krause, KH. Voltage-gated proton channels as novel drug targets: from NADPH oxidase regulation to sperm biology. Antioxid Redox Sign. (2015) 23:490–513.
doi: 10.1089/ars.2013.5806 PubMed Abstract | Crossref Full Text | Google Scholar
344. Sesti, F, Liu, S, and Cai, SQ. Oxidation of potassium channels by ROS: a general mechanism of aging and neurodegeneration? Trends Cell Biol. (2010) 20:45–51. doi: 10.1016/j.tcb.2009.09.008
PubMed Abstract | Crossref Full Text | Google Scholar
345. Panagopoulos, DJ, and Margaritis, LH. Theoretical considerations for the biological effects of electromagnetic fields In: P Stavroulakis, editor. Biological effects of electromagnetic fields : Springer (2003). 5–33. Google Scholar
346. Hall, EJ, and Giaccia, AJ. Radiobiology for the radiologist. Philadelphia: Lippincott Williams & Wilkins (2006). Google Scholar
347. Nias, AHW. An introduction to radiobiology. Chichester: J. Wiley & Sons (1998). Google Scholar
348. Reisz, JA, Bansal, N, Qian, J, Zhao, W, and Furdui, CM. Effects of ionizing radiation on biological molecules—mechanisms of damage and emerging methods of detection. Antioxid Redox Signal. (2014) 21:260–92. doi: 10.1089/ars.2013.5489 PubMed Abstract | Crossref Full Text | Google Scholar
349. Kiselyov, K, and Muallem, S. ROS and intracellular ion channels. Cell Calcium. (2016) 60:108–14.
doi: 10.1016/j.ceca.2016.03.004 PubMed Abstract | Crossref Full Text | Google Scholar
350. Blaustein, MP, and Lederer, WJ. Sodium/calcium exchange: its physiological implications. Physiol Rev. (1999) 79:763–854. doi: 10.1152/physrev.1999.79.3.763 Crossref Full Text | Google Scholar
351. Maliszewska-Olejniczak, K, and Bednarczyk, P. Novel insights into the role of ion channels in cellular DNA damage response. Mutation Res. (2024) 793:108488. doi: 10.1016/j.mrrev.2024.108488
PubMed Abstract | Crossref Full Text | Google Scholar
352. De Luca, C, Thai, JC, Raskovic, D, et al. Metabolic and genetic screening of electromagnetic hypersensitive subjects as a feasible tool for diagnostics and intervention. Mediat Inflamm. (2014) 2014:924184 Google Scholar
353. Baek, S, Quan, X, Kim, S, Lengner, C, Park, JK, and Kim, J. Electromagnetic fields mediate efficient cell reprogramming into a pluripotent state. ACS Nano. (2014) 8:10125–38. doi: 10.1021/nn502923s
PubMed Abstract | Crossref Full Text | Google Scholar
354. Giorgi, G, and Del Re, B. Epigenetic dysregulation in various types of cells exposed to extremely low-frequency magnetic fields. Cell Tissue Res. (2021) 386:1–15. doi: 10.1007/s00441-021-03489-6
PubMed Abstract | Crossref Full Text | Google Scholar
355. Liu, Y, Liu, WB, Liu, KJ, Ao, L, Zhong, JL, Cao, J, et al. Effect of 50 Hz extremely low frequency electromagnetic fields on the DNA methylation and DNA methyltransferases in mouse spermatocyte-derived cell line GC-2. Biomed Res Int. (2015) 2015:237183:1–10. doi: 10.1155/2015/237183
PubMed Abstract | Crossref Full Text | Google Scholar
356. Kaur, RP, Kaur, P, and Munshi, A. Epigenetic instability caused by oxidative stress triggers tumorigenesis In: S Chakraborti, BK Ray, and S Roychoudhury, editors. Handbook of oxidative stress in cancer: Mechanistic aspects. Singapore: Springer (2022). 1639–55. Google Scholar
357. Nishida, N, and Kudo, M. Oxidative stress and epigenetic instability in human hepato-carcinogenesis. Dig Dis. (2013) 31:447–53. doi: 10.1159/000355243
PubMed Abstract | Crossref Full Text | Google Scholar
358. Chalidis, B, Sachinis, N, Assiotis, A, Maccauro, G, and Graziani, F. Stimulation of bone formation and fracture healing with pulsed electromagnetic fields: biologic responses and clinical implications. Int J Immunopathol Pharmacol. (2011) 24:17–20. doi: 10.1177/03946320110241S204
PubMed Abstract | Crossref Full Text | Google Scholar
359. Daish, C, Blanchard, R, Fox, K, Pivonka, P, and Pirogova, E. The application of pulsed electromagnetic fields (PEMFs) for bone fracture repair: past and perspective findings. Ann Biomed Eng. (2018) 46:525–42. doi: 10.1007/s10439-018-1982-1 PubMed Abstract | Crossref Full Text | Google Scholar
360. Pilla, AA. State of the art in electromagnetic therapeutics In: M Blank, editor. Electricity and magnetism in biology and medicine. San Francisco: San Francisco Press Inc (1993). 17–22.
Google Scholar
361. Pilla, AA. Weak time-varying and static magnetic fields: from mechanisms to therapeutic applications In: P Stavroulakis, editor. Biological effects of electromagnetic fields. Berlin: Springer (2003). 34–75.
Google Scholar
362. Ryabi, JT. Clinical effects of electromagnetic fields on fracture healing. Clin Orthop Relat Res. (1998) 355S:S205–15. doi: 10.1097/00003086-199810001-00021
PubMed Abstract | Crossref Full Text | Google Scholar
363. Wade, B. A review of pulsed electromagnetic field (PEMF) mechanisms at a cellular level: a rationale for clinical use. Am J Health Res. (2013) 1:51–5. doi: 10.11648/j.ajhr.20130103.13
Crossref Full Text | Google Scholar
364. Funk, RH. Coupling of pulsed electromagnetic fields (PEMF) therapy to molecular grounds of the cell. Am J Transl Res. (2018) 10:1260–72. PubMed Abstract | Google Scholar
365. Maziarz, A, Kocan, B, Bester, M, Budzik, S, Cholewa, M, Ochiya, T, et al. How electromagnetic fields can influence adult stem cells: positive and negative impacts. Stem Cell Res Ther. (2016) 7:1–12.
doi: 10.1186/s13287-016-0312-5 PubMed Abstract | Crossref Full Text | Google Scholar
366. Pantelis, P, Theocharous, G, Veroutis, D, Vagena, IA, Polyzou, A, Thanos, DF, et al. Pulsed electromagnetic fields (PEMFs) trigger cell death and senescence in Cancer cells. Int J Mol Sci. (2024) 25:2473. doi: 10.3390/ijms25052473
PubMed Abstract | Crossref Full Text | Google Scholar
367. Meijer, DK, and Geesink, HJ. Favourable and unfavourable EMF frequency patterns in cancer: perspectives for improved therapy and prevention. J Cancer Ther. (2018) 9:188–230.
doi: 10.4236/jct.2018.93019 Crossref Full Text | Google Scholar
368. Panagopoulos, DJ, and Karabarbounis, A. Comments on behavior of charged particles in a biological cell exposed to AC-DC electromagnetic fields, and on comparison between two models of interaction between electric and magnetic fields and proteins in cell membranes. Environ Eng Sci. (2011) 28:749–51. doi: 10.1089/ees.2011.2810.com Crossref Full Text | Google Scholar
369. Panagopoulos, DJ, and Chrousos, GP. Shielding methods and products against man-made electromagnetic fields: protection versus risk. Sci Total Environ. (2019) 667:255–62.
doi: 10.1016/j.scitotenv.2019.02.344
PubMed Abstract | Crossref Full Text | Google Scholar
370. Persinger, MA. Brain electromagnetic activity and lightning: potentially congruent scale – invariant quantitative properties. Front Integr Neurosci. (2012) 6:1–7. doi: 10.3389/fnint.2012.00019
PubMed Abstract | Crossref Full Text | Google Scholar
371. Persinger, MA. Schumann resonance frequencies found within quantitative electro-encephalographic activity: implications for earth-brain interactions. Int Lett Chem Phys Astron. (2014) 30:24–32.
doi: 10.56431/p-ly2br0 Crossref Full Text | Google Scholar
372. Yan, X, Liu, X, Zhang, S, Liu, Z, and Ren, L. Study of the inhibition of Schumann resonance-inspired electromagnetic field on cancer cell proliferation. J Bionic Eng. (2025) 22:341–53. Google Scholar
373. Zahumenska, R, Badurova, B, Pavelek, M, Sojka, P, and Pavlisova, T. Comparison of pulsed and continuous electromagnetic field generated by WPT system on human dermal and neural cells. Scientific Reports. (2024) 14:5514. Google Scholar